【2.7.3】骆驼免疫球蛋白和纳米抗体(nanobody,Nb)技术
众所周知,所有骆驼科动物的血液中都有独特的抗体。与其他物种的抗体不同,这些特殊抗体不含轻链,由重链同型二聚体组成。这些所谓的重链抗体(HCAb)在VSDSJ重排后表达,需要专用的恒定γ基因。按照经典的免疫方案,在这些所谓的重链抗体中会产生免疫应答。这些HCAb易于从血清中纯化,并且抗原结合片段与靶标的相互作用较弱,这些部分对常规抗体的抗原性较低。由于单峰骆驼HCAb的抗原结合位点包含在一个单一域中,称为HCAb重链可变区(VHH)或纳米抗体(Nb),因此我们设计了一种策略来克隆免疫单峰骆驼的Nb组成选择对我们的靶抗原具有特异性的Nb。单克隆Nb在细菌中产生良好,非常稳定且高度可溶,并以高亲和力和特异性结合其同源抗原。我们已经成功地开发了用于研究目的的重组Nb,作为生物传感器的探针,用于诊断感染以及治疗癌症或锥虫病等疾病。
一、骆驼科(camelids)动物中存在重链抗体
整个哺乳动物的抗体由两条相同的H链和两条相同的L链组成(图1A)。来自骆驼科物种(即骆驼,骆驼,骆驼,骆马,鸟粪,骆驼和骆马,Camelus dromedarius, Camelus bactrianus, Lama glama, Lama guanoco, Lama alpaca and Lama vicugna)的IgG抗体是该范例的一个令人惊讶的例外,因为它们的血清中还包含相当一部分的重链抗体(HCAb) ,缺少L链(Hamers-Casterman等,1993)。 HCAb中的H链由三个而不是四个球状结构域组成(图1A)。这两个恒定结构域与经典抗体的Fc结构域(CH2SCH3)高度同源(Nguyen等,1999; Woolven等,1999)。 HCAb中缺少与经典抗体的CH1结构域相对应的结构域。因此,经典抗体Fab的抗原结合片段被还原为HCAb中的单个可变域(图1A)。在没有可变轻(VL)链结构域的情况下,这种称为VHH的可变结构域可以在抗原结合中发挥功能(uyldermans等,1994; Vu等,1997; Harmsen等,2001; aass等(2007)。反复证明,在细菌中克隆和表达的VHH是严格的单体单结构域抗原结合实体(uyldermans和Lauwereys,1999)。围绕纳米技术的宣传以及VHH的nm范围(~2.5 nm的直径和~4 nm的高度)内的扁长形状(即橄榄球球)刺激了Ablynx S,该公司专注于骆驼科抗体S的治疗应用将VHH重命名为纳米抗体(Nbs,nanobodies)。

在经典抗体的H链可变域和HCAb的可变域之间,VH和VHH分别非常相似。尽管如此,VHH和VHH包含微小但重要的差异,这些差异解释了VHHY以单结构域形式的抗原结合能力(Conrath等,2003)。两个结构域均由四个保守序列延伸(stretches),构架区,围绕(surrounding)三个高变区,互补决定区(CDR)组成(图1B)。 VH和VHH结构域采用两个β-折叠的Ig折叠,四个strands之一,五个strands之一(Muyldermans等,2001; Padlan,1994)。在此折叠中,聚集在结构域一端的三个高变区位于连接β-strand的环中,它们参与抗原识别。
- VH和VHH氨基酸序列的比对立即表明,VHH的第一个和第三个高变区比VHH的延伸范围更大,并且在单峰骆驼中但在美洲驼中,CDR1和CDR3通常包含Cys(Conrath等人(2003年)。从VHH的晶体结构中,我们知道这些Cys形成了一个二硫键,可能有助于塑造环结构(De Genst等,2006b; uyldermans等,2001)。
- VH和VHH之间的另一个明显区别出现在framework-2区域。该区域通常在VH结构域中高度保守,并由充当VL结构域锚定位置的疏水氨基酸组成。相反,VHH中的相应区域包含更多的亲水性氨基酸(图1B),这说明了VL缔合的缺失以及VHH作为单域实体的可溶行为(Nguyen等,2001)。
- 由于在VL结构域中存在三个CDR,预期经典抗体的互补位比VHH大得多。然而,已经提出通过CDR1环的延伸来扩大VHH paratope(图1B)(Nguyen等,2000; Vu等,1997)。同样,单峰骆驼的VHH中的CDR3环(在美洲驼中较少),比人或小鼠的VH中的CDR3环(平均16个氨基酸)长(分别为14或12)(uyldermans等,1994; Vu等) (1997; Wu等,1993)。显然,更长的环长度(具有更多的氨基酸)会增加互补位的大小(图1B)。
经典抗体中的许多互补位多样性源自VH-VL配对。由于在HCAbs中没有VL结构域,因此VHH的互补位显然不存在这种组合多样化。互补位库的这种限制被采用新构象的抗原结合环所补偿,这种构象在人或小鼠VH中从未观察到。实际上,VHHs的晶体结构表明,VHHs的抗原结合环不遵循规范的结构限制,显然在VHH中比在VHs中可以发现更多的环结构(Decanniere等,2000; Nguyen等。 。,2000,2001)。引入这种环多样化的原理,特别是在CDR1中,以一种微妙而巧妙的方式嵌入种系序列中。通过在有害于loop组织的位置上将两个Phe密码子(TTY)替换为Tyr密码子(TAY),VHH中的CDR1为体细胞超突变获得了两个额外的热点(TA二核苷酸)(Conrath等,2003) ; Nguyen等,2000)。实际上,CDR1中这些位置的氨基酸在VHH中的保守性比在VH中低得多。可能必须限制CDR1和CDR2对抗原识别的影响,因为对与溶菌酶复合的单峰VHHY的晶体结构调查表明CDR3是抗原结合的主要贡献者(De Genst等,2006b)。 VH-VL对的六个环或多或少地相等地对抗原结合做出贡献,而在VHH中,CDR3环通过提供至少60-80%的抗原接触而占主导地位。以前,我们注意到VHH可能在免疫过程中的亲和力成熟过程中积累了许多令人惊讶地收敛的体细胞超突变(De Genst等,2004,2005)。
除了抗原结合环结构的多样性增加而产生的互补位库增加外,单结构域形式在抗原上还留下了其互补位的足迹,其大小仅为经典抗体二结构域(VH-VL )形式的一半。与形成凹面或平坦表面的VH-VL互补位相反,VHH互补位通常由凸出的较长CDR3环(De Genst等,2006b; Desmyter等,1996)帮助形成,也是凸形的。因此,毫不奇怪地发现,单峰骆驼VHHYs优先靶向具有凹形拓扑结构的抗原表面,例如形成酶活性位点的裂口(Lauwereys等,1998)。观察到VHH经常充当酶抑制剂(或激活剂)(Lauwereys等人,1998; Transue等人,1998; Desmyter等人。 ,2002; Saerens等,2004),这一发现与美洲驼HCAb矛盾(Ferrari等,2007)。 铁汉 13:40:02
二、HCAb generation
从与人或小鼠IgH染色体基因座的同源性可以推断出,单峰骆驼基因组包含VH和VHH种系基因簇,然后是D基因库,J基因库和C基因(包括μ,γ,ε,α)。 Cδ基因在美洲驼或单峰骆驼的IgH基因座中的存在是不确定的,因为在长齿动物(猪)中也存在很长一段时间(Zhao et al。,2002)。然而,很明显,单峰骆驼的基因组包含多个Cγ基因,其中一些致力于构成HCAbs H链的保守部分,而其他Cγ基因被用于产生经典Abs的H链(Nguyen等(2003; Zou等,2005)。单峰骆驼的基因组包含约50 VH和40 VHH种系基因。 VHH种系基因的Cys编码在CDR1区域,VHH-framework-2区域内具有特征性的氨基酸取代(Nguyen等,1998)。 D基因的确切数目尚不清楚,但是,J簇似乎包含5-6基因(最上游(位于5’端)基因可能是假基因)(De Genst等人,2006a)。由于可以将相同的D和J基因重排至VH之一或VHH种系基因之一,因此可以得出结论,VH和VHH是同一基因座的一部分是很安全的(De Genst等人,2006a; Nguyen等人,2000)。在VDJ重排之后,对于VHH,似乎将Cys以体细胞形式引入CDR3区域中,但对于VH结构域而言,并不是这样。由于很难想象这是如何在DNA重组过程中完成的,因此很可能是后续选择的结果,即CDR3中没有Cys密码子的重排VHH区无法在B细胞选择过程中幸存下来,或者需要敲除CDR1中的Cys密码子缺失
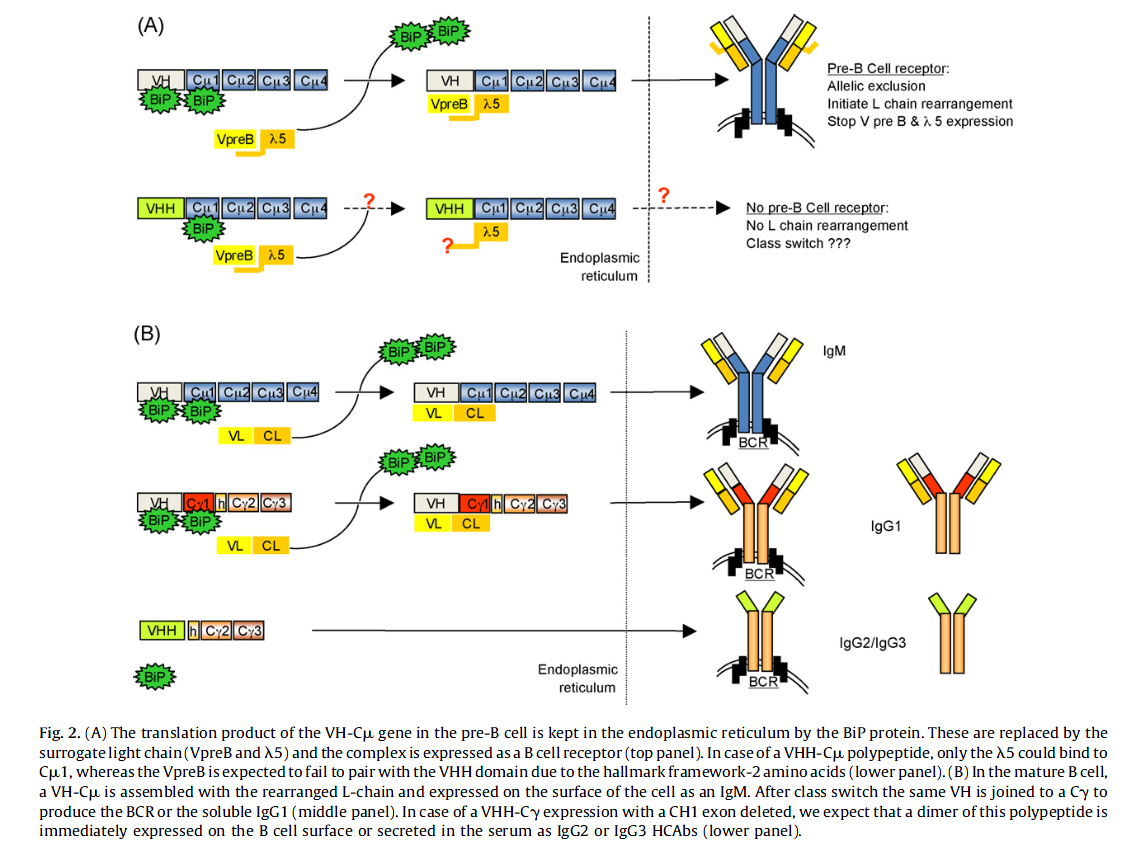
我们目前有关人类或小鼠B细胞个体发育和成熟的许多知识都来自转基因小鼠模型。但是,在B细胞产生真正的HCAb的个体发育的情况下,来自转基因小鼠的数据甚至产生了相互矛盾的结果(Bruggemann等,2006)。 HCAb专用的Cγ2a基因产生了一条H链转录本,其中的CH1外显子被删除了(因此,H链多肽中的CH1域也被删除了)。内含子5’剪接位点上保守的GT发生了单点突变。 AT负责这种特殊的VHH铰链剪接(Zou et al。,2005)。相反,在剪接位点携带具有相同突变的人Cγ基因的转基因小鼠未能从转录本中除去CH1外显子(Janssens (2006年等),目前的知识预测,在B前细胞阶段,重排的H链通过结合在V和CH1域上的BiP伴侣蛋白表达并保留在内质网(ER)中。每个H链的BiP伴侣需要被替代的L链复合物替代,以暴露前B细胞受体在细胞表面上,触发等位基因排斥,启动L链重排并终止VpreB和γ5替代L链的表达组件(artensson et al。,2007)(图2)。功能性替代L链复合物由VpreB-和γ5相关的多肽组成,其中VpreB充当VL替代物,而γ5充当CL替代物。根据我们未公开的数据,重排的VHH-DJ域用包含CH1域的CuH链表达。很难想象如何将这个附着在Cm1结构域并保留在ER中的H链的单一BiP替换为替代的L链复合物以分泌VHH-pre-B细胞受体(图2A)。我们目前的工作假设是,VHH-DJ不能与Cu结合形成pre-B细胞受体,因此无法给出启动L链重排的信号。通常,这种B细胞会进入凋亡状态。如果是HCAb个体发育,则可能尝试从Cu到Cγ类别转换,以使B细胞存活。无论如何,VHH-DJ重排的基因已在单峰H基因座中从Cu转换为Cγ基因之一,且GT转变为AT突变后,显然不再存在产生HCAb的问题(图2B)。 VHH结构域不应被BiP捕获,因为HCAb的专用Cγ基因中没有CH1结构域,原因是CH1外显子区域的3’末端共有剪接信号处存在此点突变(Elkabetz等,2005; Nguyen等人,1999; Woolven等人,1999)。
三、Nbs的应用。
基因工程技术允许快速克隆和选择抗原特异性VHHY(称为Nb)。从理论上讲,Nb可以选自大量的,合成的或naive VHH文库(Tanha等,2001; Verheesen等,2006; Yau等,2003),尽管它们通常以低亲和力识别抗原。因此,我们选择事先对单峰骆驼或骆驼进行免疫接种,因为这会给我们提供亲和力成熟的Nbs,就像在其他任何哺乳动物(如山羊和兔子)中一样,在有佐剂的情况下通过反复皮下注射对骆驼或骆驼进行免疫。经过短暂的免疫步骤(6-7周),我们从淋巴结活检或50-100-ml外周血样本中纯化淋巴细胞,RT-PCR之后,我们从样本中的B淋巴细胞的噬菌体展示载体中克隆VHH组成成分( Nguyen et al。,2001; Saerens et al。,2004)。VHH在噬菌体颗粒的顶端表达,经过2-3轮淘选后,我们可以鉴定出产生带有抗原特异性VHH的病毒体的单个克隆。从immune库中鉴定Nb很重要比从免疫scFv或Fab文库鉴定抗原结合物的相应方法更有效。在经典抗体的情况下,B细胞受体(BCR)的VH-VL对成熟为VH-VL异二聚体。在克隆互补位的结构域期间,通过PCR分别扩增VH和VL并在以后的阶段连接。这使得对于10^5个不同的BCR,需要扩增10^5个不同的VH基因区域和10^5个不同的VL基因区域,之后必须将其连接到10^10个克隆中,以涵盖从中检索作为实体成熟的原始VH-VL对的整个库。相反,对于10^5个不同B细胞的HCAb型BCR,仅当每个VHH基因包含在一个350 bp的单个基因片段中时,才需要克隆10^5个VHH的库。从此类(相对)小的文库中检索抗原特异性VHH显然比从大型文库中检索要简单得多。
一旦鉴定出具有抗原特异性VHH的克隆,通常即可在细菌系统中直接表达毫克量的可溶性,正确折叠的Nb,而酵母产生的量甚至更大(Frenken等,2000; van der Linden等,2000)。例如通常,scFv比Nbs的表达水平低10倍。此外,无论是否保质,考虑到热稳定性,抗化学变性剂抗折叠的稳定性或耐蛋白酶性,与scFv相比,Nbs的稳定性都更高(Dumoulin等,2002; Harmsen等。 ,2006)。 Ablynx(www.ablynx.com)报道了功能形式的Nb成功通过肠道。尽管Nb的特异性和亲和力可与更好的scFv或Fabs轻松竞争,但严格的单体行为及其仅350 bp的小基因使Nb理想地适用于较大的二价或双特异性构建体(Conrath等,2001)。
在我们的实验室以及其他实验室中,Nb已被用作研究工具,并用于各种诊断或治疗应用。为了增加VHH的多功能性,已鉴定出稳定表达的稳定VHH结构域,并充当环受体以嫁接其他不稳定或表达欠佳的VHH的环,从而形成具有增强功能的嵌合域环供体VHH的稳定性,更好的表达以及抗原特异性和亲和力(Saerens等,2005)。
在另一项工作中,我们已经在Nb后面克隆了猪Fc同种型,以得到由VHH-pigFc组成的嵌合HCAb(请参阅Butler等人的文章)。该嵌合蛋白用于产生针对猪IgG同种型的单克隆抗体。按照相同的策略,用与β-内酰胺酶融合的Nb基因或人ApoLI 或mRFP(分别为载脂蛋白L-I或单体红色荧光蛋白)的截短基因版本生成遗传构建体,我们获得了用于治疗或研究的有趣构建体目的(Baral等人,2006; Cortez-Retamozo等人,2004; Rothbauer等人,2006)。内酰胺酶融合构建体包含针对人肿瘤标志物癌胚抗原(CEA)的Nb。向携带人肿瘤的冰中注入这种蛋白融合物,以使缀合物负载于肿瘤。通过内酰胺酶将在小鼠静脉中施用的前药转变成高毒性分子。因此,主要在肿瘤部位附近产生高浓度的毒素。遵循这种抗体依赖性酶前药治疗方法,我们可以彻底根除,而不会复发在小鼠模型中建立的实体瘤(Cortez-Retamozo等,2004)。
通过将截短的ApoLI的结构连接到Nb上,该Nb可以识别罗德锥虫(Trypanosome rhodesiensis),这是一种导致东非昏睡病的寄生虫,我们产生了一种强力的锥虫分解性化合物,可以消除血液中的寄生虫(Baral等人,2006; Stijlemans等人(2004年)。该Nb识别锥虫变体表面糖蛋白的保守碳水化合物,该蛋白是覆盖寄生虫整个表面的主要外壳蛋白,在抗原变异过程中被其变体之一交换,这是逃避宿主免疫应答的主要寄生虫机制。 。最后,用mRFP(称为染色体抗体)扩展Nb产生了靶向和揭示抗原在活细胞中的驻留的有用工具。用针对细胞角蛋白和层粘连蛋白的Nb进行了原理验证(Rothbauer等,2006)。在共聚焦显微镜下揭示了用相应的Nb-mRFP的基因构建体瞬时转染的细胞系,这令人信服,图片中的抗原被染色体染色。此外,近来已证明,即使以低剂量存在于复杂混合物中,拴在固相支持物上的Nb也是捕获其同源抗原的理想选择(Rothbauer等,2008)。因此,我们可以将Nb推荐为一种高度抗原特异性的免疫沉淀工具,尽管在Western blot中其性能相当差。总之,在用抗原对美洲驼,骆驼或单峰骆驼进行短时间免疫后,从免疫动物体内克隆VHH组成成分并进行噬菌体展示,即可轻松鉴定出抗原特异性Nb。 Nb在微生物系统中的良好表达和有益的生化特性(良好的溶解性,在恶劣条件下的稳定性,对抗原的高亲和力和特异性,小尺寸和严格的单体行为)使Nbs成为用于研究目的或诊断或治疗应用的理想工具。
参考资料
- Veterinary Immunology and Immunopathology Volume 128, Issues 1–3, 15 March 2009, Pages 178-183. Camelid immunoglobulins and nanobody technology
