【8.1.3】个性化新抗原肿瘤疫苗的研究进展和挑战
免疫系统识别并消除癌细胞的过程是复杂的,并受许多因素的调控,这些因素中就包括癌细胞中突变发生的质量和数量。其中最重要的过程之一就是肿瘤新抗原的产生。肿瘤新抗原是呈现在肿瘤细胞表面上的肽,与主要组织相容性复合物(MHC)分子相结合,以蛋白质复合的形式存在于肿瘤细胞表面,可被细胞毒性T细胞受体(TCR)特异性识别,从而激活T细胞的免疫反应。从免疫学的角度来看,肿瘤新抗原是真正的外源蛋白,是在正常人体器官/组织中是完全不存在的。
对于人类大多数没有病毒病因的肿瘤,肿瘤新抗原可能来自
- 多种非同义的遗传改变,包括单核苷酸变异(SNVs)、插入和缺失(InDels)、基因融合、移码突变和结构变异(SV)。
- 对于与病毒相关的肿瘤,如与HPV相关的宫颈癌或口咽癌,与MCV相关的默克尔细胞癌(MCC)和EBV相关的头颈癌,从病毒基因组中开放阅读框(ORF)衍生的任何表位也是新抗原的潜在来源。
近年来,随着二代测序技术和生物信息技术的进步,肿瘤精准免疫治疗得到来了很大的发展,肿瘤新抗原的研究更是为肿瘤免疫治疗提供了新的研究方向。癌症免疫疗法包括:
- 肿瘤疫苗接种
- 过继性免疫细胞治疗 [ACT,包括了TIL(肿瘤浸润性淋巴细胞治疗)、TCR-T(T细胞受体嵌合型T细胞)、CAR-T(嵌合抗原受体T细胞)
- 免疫检查点阻断(如使用PD-1/PD-L1和CTLA-4抗体)
- 非特异性免疫刺激(包括淋巴因子激活的杀伤细胞疗法、细胞因子介导的杀伤细胞疗法)。
从理论上讲,新抗原是理想的免疫治疗靶标,因为它们可以被宿主免疫系统识别为非自身抗原。因此,新抗原特异性免疫反应不容易受到复杂的免疫耐受机制的影响。另外,新抗原应该不太可能触发自身免疫反应,因为它们在正常细胞上不表达。也正因为如此,新抗原成为肿瘤疫苗以及以T细胞为基础的癌症免疫疗法中及具吸引力的靶标。基于新抗原的个体化肿瘤疫苗和过继性T细胞免疫治疗已显示出令人鼓舞的初步结果。此外,最近的研究表明,新抗原在免疫逃逸,免疫编辑和对免疫检查点抑制剂的敏感性中具有重要作用。
一、临床前及临床研究中的肿瘤新抗原应用
越来越多的证据表明,新抗原在以肿瘤特异性T细胞介导的抗肿瘤免疫治疗中起着关键作用:
- 通过新抗原的T细胞识别诱导抗肿瘤免疫反应;
- 肿瘤突变/新抗原负荷与免疫检查点阻断的临床结果之间存在关联;
- 基于新抗原的肿瘤疫苗或过继性T细胞免疫治疗显示出令人鼓舞的初步抗肿瘤作用。
二、新抗原用于个体化肿瘤疫苗的开发
靶向新抗原的个体化疫苗旨在在体内诱导和扩增新抗原特异性T细胞群体,以增强个体之间的过继性抗肿瘤免疫力。实际上,肿瘤疫苗首先被用于靶向TAA(肿瘤相关抗原),TAA在肿瘤中过表达,但也在正常组织中表达。因此,由于TME(肿瘤微环境)中的T细胞活化不足,既往靶向TAA的肿瘤疫苗的临床试验未能证明持久有效的治疗功效。相比之下,通过NGS或质谱检测到的新抗原可导致宿主免疫系统的免疫耐受性降低和安全性提高。
近期临床前研究表明,靶向新抗原的肿瘤疫苗在黑色素瘤、结肠癌、食道鳞状细胞癌、肉瘤和神经胶质瘤的鼠类肿瘤模型中具有有效性和可行性。Theresa等人合成了含有IDH1(R132H)p123-142突变区的新肽,接种到神经胶质瘤的小鼠模型中,疫苗接种后的结果表明,新肽疫苗可在具有IDH1 R132H突变的神经胶质瘤小鼠模型中产生有效的突变特异性抗肿瘤免疫反应。临床前的研究也加快了新抗原肿瘤疫苗的临床试验的发展。Beatriz的团队于2015年首次报道了新抗原疫苗的I期临床试验。他们发现在三名晚期黑色素瘤患者中,树突状细胞(DC)疫苗可以促进HLA-A * 02:01呈递新抗原。在2017年发表了两项重要研究确认了基于新抗原的疫苗在黑素瘤治疗中的强效作用:
- 在6例黑素瘤患者中使用了多达20种肿瘤新抗原的合成长肽疫苗,其中有4例在接种疫苗后25个月内未复发,而另外2例在复发时接受了PD-1抗体治疗并达到了肿瘤完全反应(CR);
- 在13例黑色素瘤患者中,使用个体化的RNA突变疫苗接种联合PD-1阻断疗法,三分之一的患者实现了CR,并且体内检测到的新抗原特异性T细胞活化增强。
除了黑色素瘤,近期的临床研究也表明基于新抗原的疫苗对于胶质母细胞瘤等“冷”肿瘤也是可行的。
新抗原用于过继性T细胞转移治疗的开发 与基于新抗原的肿瘤疫苗相比,基于新抗原的ACT治疗具有若干潜在优势,这些优势包括:
- 具有更高的对新抗原起反应的T细胞群(NRTs, neoantigen-reactive T cells );
- 在NRTs扩增阶段,来自肿瘤微环境的免疫抑制作用较小。
多项研究已证实,来自免疫原性突变的新抗原可诱导肺癌、头颈部鳞状细胞癌、大肠癌、乳腺癌和淋巴瘤中新抗原特异性T细胞的活化。将新抗原特异性TCR序列转导至患者的外周血淋巴细胞(PBLs)中可能更起到抗肿瘤作用。临床前研究数据证明,与KRAS突变新肽反应的TCR工程PBLs减少了黑色素瘤和其他瘤种在小鼠模型上的异种移植,这一证据支持了TCR-T治疗的可行性。在基于新抗原的ACT疗法的临床研究中,已有成功治疗的先例。Rosenberg的研究团队首先通过过继转移CD4+ TILs治疗转移性上皮癌患者,并在2014年实现肿瘤消退。通过将特异性TCR序列转导至特定的CD8+ T细胞中,可以为具有相同突变的患者生产新的抗原特异性TCR-T细胞,从而扩大和增强新抗原反应性TILs的治疗应用和效果。
三、开发与新抗原相关的组合疗法
由于PD-1/PD-L1通路对CD8+ T细胞介导的抗肿瘤免疫具有免疫抑制作用,新抗原疫苗和ACT治疗加上抗PD-1/PD-L1抗体,可以产生强大的抗肿瘤免疫应答。在初步研究中,两个研究小组独立地证实,PD-1/PD-L1阻断能显著改善新抗原疫苗诱导的免疫应答。从理论上讲,一些癌症常规疗法可能导致肿瘤相关抗原和新抗原的释放,这表明常规疗法与新抗原疫苗疗法可能具有协同作用。一些化疗药物已显示出通过改变新抗原组库、增加抗原产生、改善抗原呈递以及增强T细胞的运输和反应来增强肿瘤细胞的抗原性和免疫原性。或许,我们不应该简单地将化疗看作是抑制肿瘤的一种疗法,而应将其视为宿主免疫系统的积极调节方式。
四、新抗原可作为免疫检查点抑制剂的潜在生物标志物
对于高肿瘤突变负荷(TMB)或肿瘤新抗原负荷(TNB)的肿瘤,免疫检查点阻断已显示出明显的治疗反应。一系列临床试验显示,NSCLC或黑色素瘤患者对免疫检查点阻断具有较高的客观反应。在实体瘤中,这两种肿瘤类型具有最高的TMB。相反,具有低TMB/TNB的癌症(如前列腺癌)几乎不能从免疫检查点抑制剂中获益。这些结果为新抗原在抗肿瘤免疫反应中的重要性提供了补充依据。当然,这些结果不能反应TNB与免疫检查点抑制剂之间的线性关联。如,一些TNB高的患者对免疫检查点阻断疗法无反应,而另一些TNB低的患者则易受免疫检查点抑制剂的影响。高TNB仅代表存在更高免疫原性新抗原的可能性。当把MHC/HLA呈递新抗原的可能性和新抗原被T细胞识别纳入新抗原计算模型中时,观察到该类模型可以更好地预测抗CTLA-4治疗黑色素瘤和抗PD-1治疗肺癌的生存情况。有研究报道了新抗原异质性在抗肿瘤免疫中具有重要作用。
新抗原异质性可能是免疫检查点抑制剂的耐药机制之一 基于肿瘤新抗原在免疫检查点抑制剂应答中的重要作用,可以合理地假设新抗原克隆演变将导致免疫检查点阻断的耐药性。一项回顾性研究纳入了42例接受PD-1抗体单药治疗或PD-1+ CTLA-4联合阻断治疗的NSCLC患者,并研究了获得性耐药出现期间不断发展的新抗原演变。在四例获得性耐药患者中,发现由消除肿瘤亚克隆或删除含有截短突变的染色体区域而导致的新抗原丢失与T细胞受体克隆性的改变有关,从而导致了对免疫检查点阻断的获得性耐药。这些结果暗示新抗原丢失可能是潜在的耐药机制之一。扩大新抗原反应性的广度可能会延迟免疫检查点阻断获得性耐药的发展。
因此,准确、快速并且经济有效的鉴定新抗原对免疫治疗的成功至关重要。
五、肿瘤新抗原的来源
当前的免疫疗法中对新抗原的研究主要集中在体细胞突变上,并且这种类型的新抗原与免疫疗法之间的相关性已被广泛报道。NGS技术的应用为肿瘤突变的各种来源提供了进一步的证据支持。已发现源自这些突变的新抗原能够引起CD8或CD4淋巴细胞的有效应答,这些新抗原可用作肿瘤疫苗或ACT免疫疗法的治疗靶点。目前发现的各种类型的新抗原不只来自于基因组水平的突变,还包括来自于转录组水平的突变和蛋白质组学水平的替代(图1)。
总体来说,肿瘤新抗原的来源可分为六类:
- 基因组水平的突变,包括SNVs和小片段InDels;
- 基因组水平的基因融合;
- 转录组水平的选择性剪接;
- 转录组水平的RNA编辑;
- 转录组水平的翻译蛋白质的非编码区突变;
- 蛋白质组水平的替代剪接。
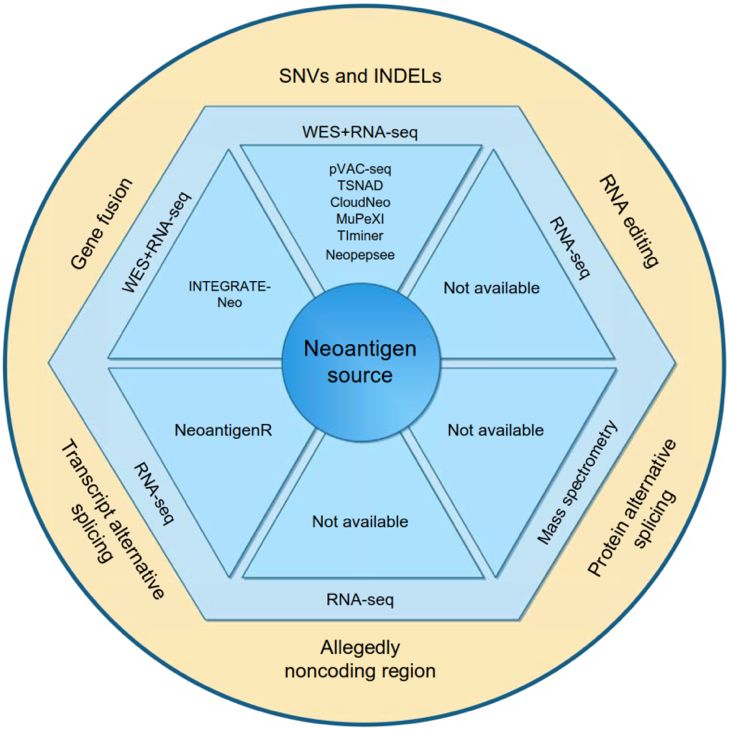
图1 各种类型新抗原的来源和可利用的计算机模拟工具
基因组水平——SNVs和小片段InDels
SNVs是肿瘤中最常见的基因组水平的突变类型,编码蛋白质的非同义突变产生的突变肽很可能会被MHC-I呈递到肿瘤细胞表面。不同瘤种之间的突变频率变化范围超过1000倍。许多研究集中在基于SNVs的新抗原上。Charoentong等人在TCGA数据库中,对20种实体瘤的肿瘤内免疫全景和肿瘤新抗原进行了表征,并创建了“癌症免疫组图谱”(The Cancer Immunome Atlas)。此外,一些研究将多种类型癌症患者的体系突变负荷和基于SNV的新抗原负荷与ICB治疗的临床获益联系起来。InDels可产生新的开放阅读框(ORF),以及大量非自身肽,也有助于产生免疫原性表型。对来自TCGA的19种癌症类型的5777例实体瘤进行的大规模分析表明,由InDel产生的新抗原比由SNV产生的新抗原更具免疫原性。同样,由InDel产生的新抗原负荷也与黑色素瘤的ICB应答显著相关。已经证明,由SNV和InDel产生的新抗原均可被MHC-I和MHC-II呈递,并能充分引起CD8 +和CD4 + T细胞免疫反应。这两种类型的基因组突变是基因组水平上肿瘤新抗原的主要来源。
基因组水平——基因融合
大多数融合基因会产生框外转录本,这些转录本可能会产生较长的新的蛋白序列和大量的候选新抗原。Gao等人通过分析TCGA中具有33种癌症类型的9624个样本队列来探索基因融合全景,研究揭示了移码融合(frameshift fusions)比框融合(frame fusions)可以产生更多的免疫原性新肽。Yang等人证明基因融合是免疫原性新抗原的来源,并且报道了由MHC-1复合物呈递的DEK–AFF2和NFIB–MYB两种基因融合可引发有效的CD8 + T淋巴细胞免疫反应。Makita等人鉴定了DEK-CAN和BCR-ABL的两个基因融合事件,它们产生的新抗原可被MHC-II限制性CD4 + T淋巴细胞识别。尽管与SNV和InDel衍生的新抗原相比,基于基因融合的新抗原较少出现,但实验验证表明它们倾向于具有更高的免疫原性。
转录组水平——转录本选择性剪接
选择性剪接是一个受多种因子调节的过程。剪接因子(如U2AF1和SF3B1)的体系突变可能导致癌细胞的剪接过程出现大范围的景观变化。Kahles等人对TCGA中8705名患者的选择性剪接突变进行了泛癌分析。该研究揭示由选择性剪接产生的肿瘤特异性新连接是新抗原的重要来源。特别是内含子保留剪接(一种导致mRNA转录本中包含内含子的RNA剪接错误)是肿瘤新抗原的来源。一些研究也证实了由这种类型产生的抗原新表位可由MHC-I或MHC-II分子呈递,并引发CD4+或CD8+淋巴细胞的某种免疫反应;当然,它们对不同癌症类型的免疫原性有待系统性研究。
转录组水平——RNA编辑
RNA编辑是一种常见的转录后修饰,会改变RNA序列中的特定核苷酸,也可能导致非同义氨基酸替代,进而产生新蛋。A-to-I RNA编辑是人类最常见的RNA编辑类型,可促进包括癌症在内的多种疾病的转录组和蛋白质组学多样性。RNA编辑可通过增加自身抗原负荷来促进自身免疫性疾病系统性红斑狼疮的进展。A-to-I RNA编辑产生的肽由MHC-I分子呈递,并引发CD8 + T细胞反应,这表明RNA编辑扩展了由人类白细胞抗原(HLA)呈递的自身抗原种类,且这些肿瘤新抗原可被免疫系统识别。迄今为止,尚未报道过来自RNA编辑的可以启动CD4 + T细胞活化的MHC-II限制性新抗原。源于RNA编辑的新抗原图谱分析和免疫原性的研究正在进行,以阐明此类型转录组突变在免疫治疗中的临床意义。
转录组水平——编码蛋白的非编码区
百分之九十九的肿瘤突变发生在非编码区,外显子组仅覆盖人类基因组的2%。然而,人类高达75%的基因组可以被转录并潜在地翻译成蛋白质。因此,仅关注外显子组作为肿瘤特异性新抗原的来源是有局限的。最近的研究表明,许多所谓的非编码区实际上是蛋白质编码区,可以产生MHC-I呈递的许多肽,其中一些肽已被确定为TILs的靶标。但是,目前尚未报道可能启动CD4 + T细胞活化的MHC-II限制性新抗原来源于非编码区。与其他基因组和转录组水平的突变相比,源自非编码区的新抗原很少被提及。需要开发有效的计算工具来识别此类新抗原并对其免疫原性进行验证。
蛋白组水平——蛋白组学上的选择性剪接
蛋白酶体也可以剪切和粘贴肽序列,然后释放与原始蛋白质序列不匹配的肽。几项研究报告说,蛋白酶体产生的剪接肽具有抗原性,且可以被CD8 + T淋巴细胞识别。尽管尚未报道可启动CD4 + T细胞活化的由蛋白质组学水平上可变剪接肽衍生的MHC-II限制性新抗原。研究表明蛋白酶体产生的剪接肽占MHC-I免疫肽组总数的三分之一,这可能为肿瘤疫苗或免疫疗法提供了新的新抗原来源。
现阶段,我们对来自各种潜在来源的新抗原的鉴定,以及有关的计算问题的综合研究仍是缺乏的。那么,问题来了:
- 我们如何基于多组测序数据鉴定和区分源自各种变异产生的新抗原,并确定其优先级,并为此开发集成工具?
- 们如何开发一种有效的一站式计算管线,以鉴定从各种数据源获得的新抗原,并将其应用于大数据队列以评估新抗原对ICB治疗反应的预测能力?
- 我们如何将测序数据与HLA-肽质谱(MS)数据相结合来提高新抗原鉴定的特异性?
参考资料
- https://www.sohu.com/a/348342259_120051885
- Jiang T, Shi T, Zhang H, et al.Tumor neoantigens: from basic research to clinical applications.J Hematol Oncol. 2019 Sep 6;12(1):93.
- Zhou C, Zhu C, Liu Q.Toward in silico Identification of Tumor Neoantigens in Immunotherapy.Trends Mol Med. 2019 Sep 4. pii: S1471-4914(19)30209-6.
- Laumont CM, Vincent K, Hesnard L et al.Noncoding regions are the main source of targetable tumor-specific antigens.Sci Transl Med. 2018 Dec 5;10(470).
