【1.3.2】免疫细胞--T淋巴细胞
一、T 淋巴细胞在胸腺中的分化发育
T 淋巴细胞是骨髓淋巴样干细胞进入胸腺后经过有序分化发育形成的具有 免疫功能的成熟 T 细胞,故称胸腺依赖性淋巴细胞(thymus dependent lymphocytes),简称 T 淋巴细胞或 T 细胞。T 细胞约占血液中淋巴细胞总数的 70%~80%,在淋巴结和脾脏也大量存在。
根据 T 细胞表面分子组成和功能特性, 可将其分为 CD4+ Th1、CD4+Th2、CD4+T h17、CD4+Tfh 细胞和 CD8+CTL 细胞等亚群:
- 其中 CD4+ Th1 细胞、CD4+T h17 细胞和 CD8+CTL 细胞是执行适应性细胞免疫应答 的免疫细胞;
- CD4+Th2 细胞和 CD4+Tfh 细胞是协助 B 淋巴细胞产生适应性体液免 疫应答的免疫细胞;
- 调节性 T 细胞包括 CD4+CD25+Foxp3+ 自然调节 T 细胞和 CD4+ 诱导性调节 T 细胞,他们对初始 T 细胞、活化 CD4+/CD8+T 细胞和某些固有免疫细 胞具有抑制作用。
αβT 淋巴细胞在胸腺中的分化发育过程如图所示:骨髓淋巴样干细胞随血 液进入胸腺后,经早期发育、阳性选择和阴性选择三个阶段,分化发育为具有免 疫潜能的成熟 T 细胞,即初始 T 细胞(naive T cell)。
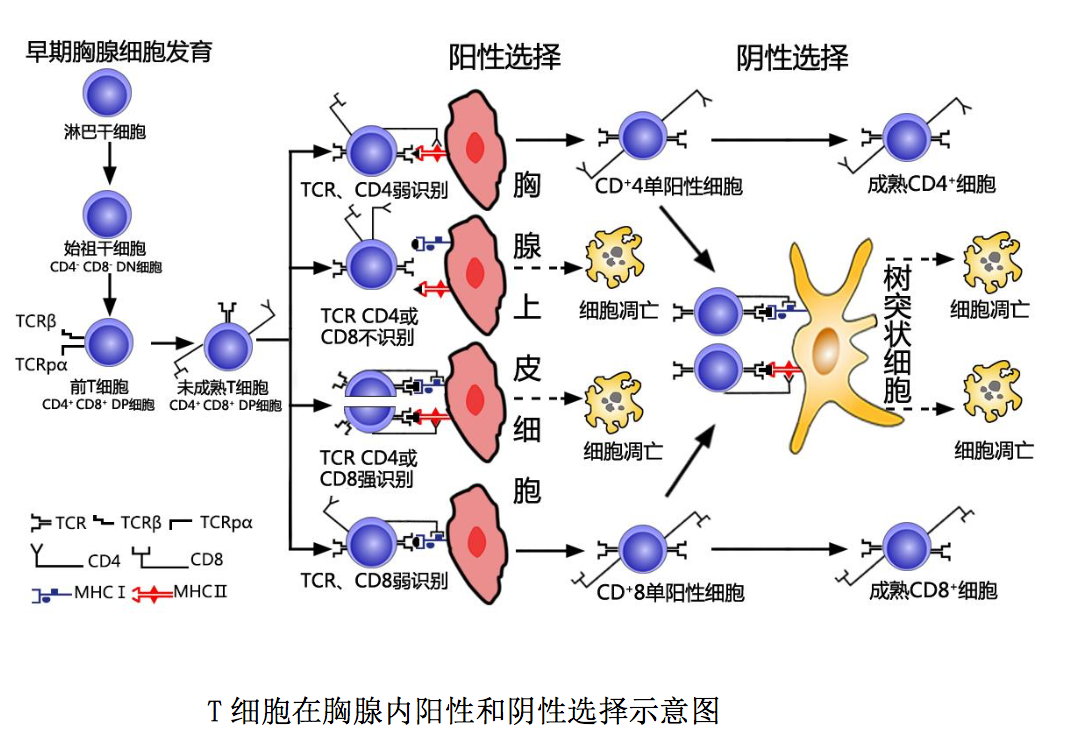
1.1 早期胸腺细胞发育阶段
淋巴样干细胞随血液进入胸腺皮质与髓质交界 处后称为始祖 T 细胞(Pro-T cell)。始祖 T 细胞不表达 T 细胞受体(TCR)和 CD3 分子,也不表达 CD4 和 CD8 分子,故称 CD4-CD8-双阴性胸腺细胞 (double negative thymocytes ) ,简称 DN 细胞。此种 DN 细胞表面具有多种胸腺激素和 IL-7 等细胞因子的受体;在胸腺微环境中胸腺激素和 IL-7 等细胞因子作用下, 可增殖分化为低表达 CD3 分子的 CD4+CD8+双阳性细胞(double positive cell )、 简称 DP 细胞,即前 T 细胞(Pre-T cell)。此种前 T 细胞 TCRβ链基因发生重排, 成功表达β链并与一条称之为前 T 细胞 α 链(pTα)的替代链配对组成前 T 细 胞受体(TCR pTαβ);后者与 CD3 分子结合形成 TCR pTαβ-CD3 复合体表达于 前 T 细胞表面。上述前 T 细胞增殖活跃、数量庞大;但只有少部分(约 5%)发生 TCRα 链基因重排,成功表达 α 链并与β链配对组成具有识别功能的 T 细胞受 体,即成为低表达 TCRαβ-CD3 复合体的 CD4+CD8+双阳性未成熟 T 细胞.
1.2 阳性选择 (positive selection) 阶段
上述双阳性未成熟 T 细胞表 面 CD4 和 CD8 分子是识别结合 MHC II类和 I 类分子的受体。在胸腺皮质区,此 种未成熟 T 细胞通过表面 CD4/CD8 分子与胸腺上皮细胞表面 MHC-II/I 类分子低 亲和力结合(弱识别),或通过表面 TCR-CD3 复合体和 CD4/CD8 分子与胸腺上皮 细胞表面自身抗原肽-MHCII/I 类分子复合物低亲和力结合(弱识别)相互作用 后,可分化发育为高表达 TCR-CD3 复合体的 CD4+或 CD8+单阳性细胞 (single positive cell),简称SP细胞;而高亲和力(强识别)或未与胸腺上皮细胞表 面自身抗原肽-MHCII/I 类分子复合物结合的双阳性未成熟 T 细胞则发生凋亡, 此即胸腺中的阳性选择过程。通过阳性选择,可获得能够识别结合自身 MHCII/I 类分子或自身抗原肽-MHCII/I 类分子复合物的 CD4+或 CD8+单阳性未成熟 T 细胞。
1.3 阴性选择(negative selection)阶段
阴性选择主要发生于胸腺皮质 与髓质交界处,位于该处的胸腺树突状细胞和巨噬细胞高表达自身抗原肽-MHC II类或 I 类分子复合物。当上述单阳性未成熟 T 细胞通过表面 TCR-CD3 复合体和 CD4/CD8 分子与胸腺树突状细胞、巨噬细胞表面相应自身抗原肽-MHCII/I 类分 子复合物高亲和力结合(强识别)相互作用后发生凋亡,从而导致体内高亲和力 自身反应性 T 细胞被清除,即对自身抗原形成中枢免疫耐受。而那些以低亲和 力或未能与树突状细胞、巨噬细胞表面自身抗原肽-MHCII/I 类分子复合物结合 的单阳性未成熟 T 细胞则得以存活,进而分化发育为 CD4+/CD8+单阳性成熟 T 细 胞,此即胸腺内的阴性选择过程。通过阴性选择分化发育成熟的未曾接受相应抗 原刺激的 CD4+/CD8+单阳性 T 细胞具有免疫潜能,称为初始 T 细胞。他们通过血 液循环进入外周免疫器官后,可接受抗原刺激、产生适应性免疫应答。
二、T 淋巴细胞表面分子及其主要作用
2.1 TCR-CD3 复合体
是 T 细胞表面 T 细胞受体(TCR)与 CD3 分子非共价结 合形成的复合体;其中 T 细胞受体(T cell receptor,TCR)是 T 细胞表面特异 性识别抗原的受体;CD3 是传递细胞活化信号的免疫分子,也是所有 T 细胞的特 征性表面标志。T 细胞表面 TCR 是由α和β两条肽链通过链间二硫键连结组成的 TCRαβ异二聚体,其结构如图 10-2 所示:TCRαβ异二聚体由胞外、跨膜和胞 内区三个部分组成,每条肽链胞外区均有两个结构域,即靠近氨基(N)端的可 变区(V 区)和靠近细胞膜的恒定区(C 区)。TCRα链和β链可变区与 Ig 可变区 非常相似,各有三个氨基酸组成和排列顺序高度易变的超变区(HVR),从而造就 了 TCRαβ的高度多样性。TCRαβ超变区是与相应抗原肽-MHC 分子复合物特异 性识别结合的部位,又称互补结合区(complementarity determining region, CDR)。TCRαβ胞内区短小,没有传递细胞活化信号的作用;其疏水性跨膜区含 带正电荷的氨基酸残基(赖氨酸或精氨酸),借此能与跨膜区带负电荷氨基酸残 基(天冬氨酸)的 CD3 多肽链非共价结合组成 TCRαβ-CD3 复合体。
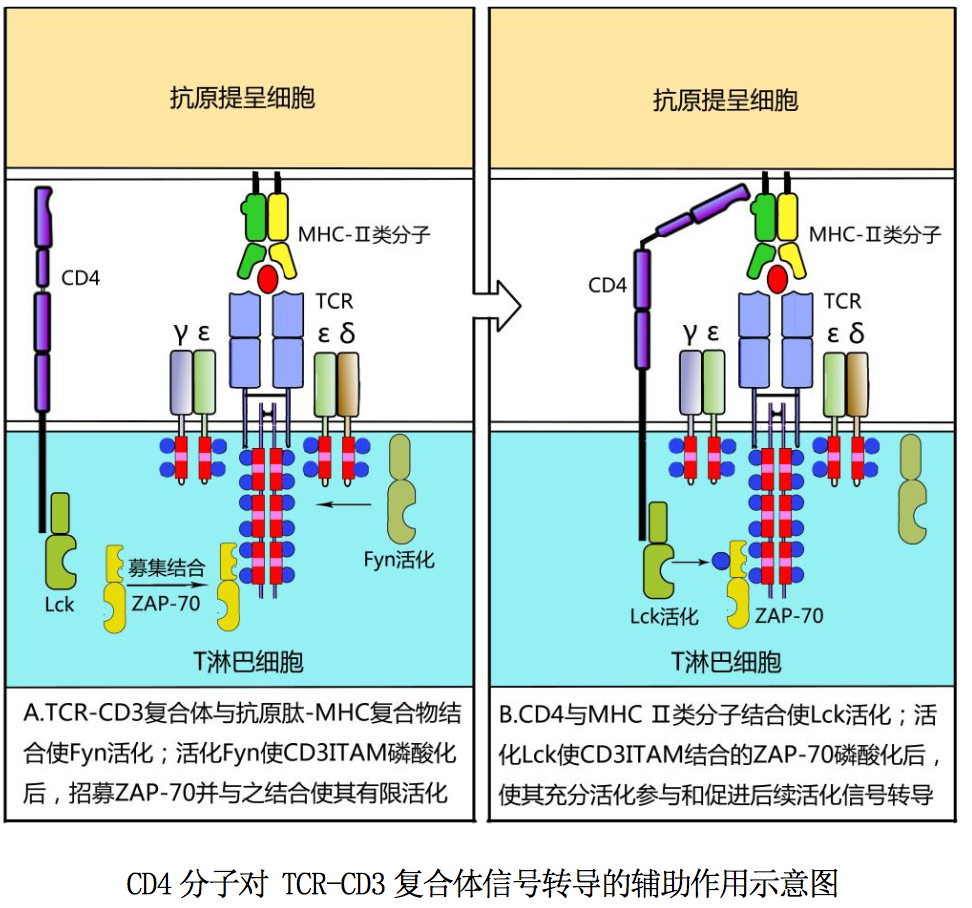
CD3 分子由γ、δ、ε、ζ和η五种肽链组成,其中ε链分别与γ链和δ链 非共价结合组成γε和δε异二聚体;ζ链多以ζζ同源二聚体形式存在,也能 以ζη异二聚体形式存在。CD3 分子如图 10-2A 所示由三对二聚体,即γε异二 聚体、δε异二聚体和ζζ同源二聚体组成:上述各条肽链跨膜区均含有带负电 荷的氨基酸残基,借此能与 TCRαβ 跨膜区带正电荷的氨基酸残基非共价结合组 成 TCR-CD3 复合体。CD3 分子胞内区均含免疫受体酪氨酸激活基序/模体(immune receptor tyrosine-based activation motif,ITAM)。ITAM 是免疫细胞激活性 受体分子胞内段所携带的结构,其内所含酪氨酸残基发生磷酸化后可招募含有SH2 结构域的蛋白激酶(如 ZAP-70)或衔接蛋白,参与启动细胞活化信号的转导。
CD3 分子介导的活化信号转导如图所示:T 细胞通过表面 TCR-CD3 复合体与 APC 表面抗原肽-MHC 分子结合相互作用,可使胞质内与 CD3ζ链尾肽相关的蛋白 酪氨酸激酶 Fyn 活化,从而导致 CD3 各条肽链胞质区 ITAM 中酪氨酸残基磷酸化; 磷酸化 ITAM 可募集、结合蛋白酪氨酸激酶 ZAP-70,为启动 T 细胞后续活化信号 转导做好准备。


2.2 TCR 辅助受体
CD4 分子和 CD8 分子是 T 细胞表面的 TCR 辅助受体。CD4 分子是一种分子量为 55kD 的跨膜糖蛋白,在细胞膜上以单体形式存在,为 Ig 超家族成员。CD4 分子是识别结合 MHCII类分子的受体,其胞外区能与 MHCII类 分子β链 Ig 样区的β2 结构域结合;其胞内区尾肽与蛋白酪氨酸激酶 Lck 相关 联,参与胞内活化信号的转导(图 10-3A)。CD4 分子也是人类免疫缺陷病毒(HIV) 壳膜蛋白 gp120 的受体,因此 HIV 可选择性感染 CD4+T 细胞,引发获得性免疫 缺陷综合征。CD8 分子是由α和β链借二硫键连接组成的跨膜糖蛋白,为 Ig 超 家族成员。CD8 分子是识别结合 MHCI类分子的受体,其胞外区能与 MHCI类分 子α链 Ig 样区的α3 结构域结合;其胞内质区尾肽与蛋白酪氨酸激酶 Lck 相关 联,参与胞内活化信号的转导。
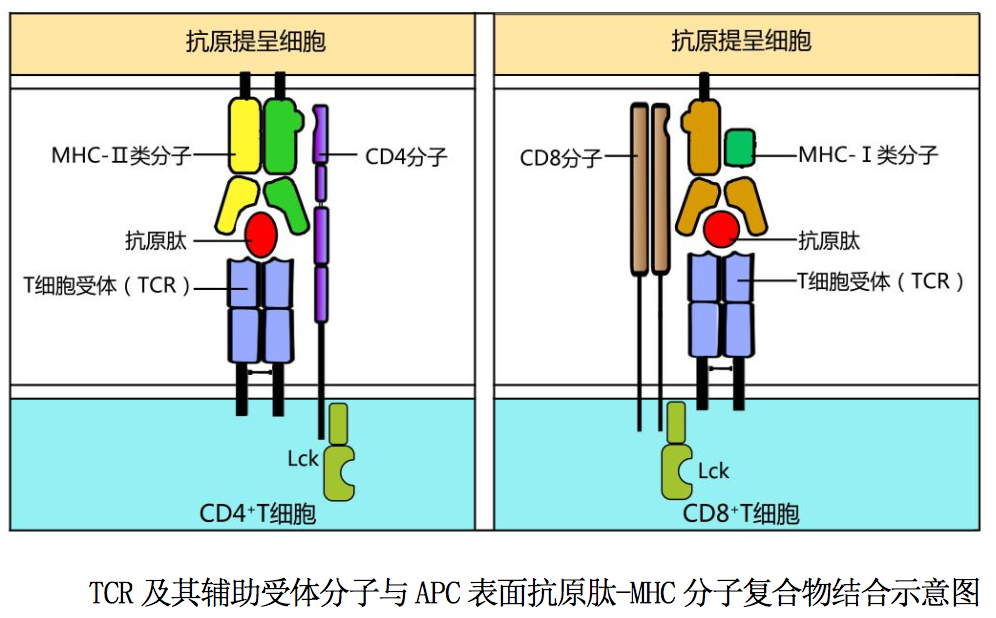
现以 CD4 分子为例,简述其对 TCR-CD3 复合体的辅助作用:CD4+T 细胞通过表面 TCR -CD3 复合体与 APC 表面相应抗原肽-MHCII类分子复合物结合,可使其表面 CD4 分子与上述抗原肽-MHCII类分子复合物中的 MHCII类分子(β2 结构域)结 合,显著增强 CD4+T 细胞与 APC 之间的相互作用;并使 CD4 分子聚集在 TCR-CD3 复合体周围,导致胞浆内与 CD4 分子尾肽相关的蛋白酪氨酸激酶 Lck 活化,后者 可促进 CD3 胞质区与 ITAM 结合的 ZAP-70 活化,从而加速 T 细胞后续活化信号的 转导。因此 CD4 分子被称为 T 细胞受体的辅助受体,简称 TCR 辅助受体。
2.3 共刺激分子(co-stimulating molecule, CM)
表达于 APC 和 T、B 淋 巴细胞表面的黏附分子,因其具有介导产生共刺激信号的作用,又称共刺激分子 或协同刺激分子。
(1)CD28:是 Ig 超家族成员,以同源二聚体形式表达于 CD4+T 细胞和半数 以上 CD8+T 细胞表面。CD28 分子是 T 细胞表面的黏附分子,也是最重要的共刺激 分子,其胞外区结构域能与 APC 表面相应共刺激分子,即 B7-1/B7-2(CD80/CD86) 互补结合;其胞内区与多种信号分子相连,参与活化信号的转导。T 细胞通过表 面 TCR-CD3 复合体和 CD4/CD8 分子与 APC 表面相应抗原肽- MHCII/I类分子复 合物结合,可诱导产生 T 细胞活化第一信号;通过其表面 CD28 分子与 APC 表面 相应配体 B7-1/2 分子结合,可诱导 T 细胞产生共刺激信号(T 细胞活化第二信 号)导致 T 细胞活化。
(2)细胞毒性 T 淋巴细胞抗原-4(cytotoxic T lymphocyte antigen-4, CTLA-4):为同源二聚体,主要表达于活化 T 细胞表面,静息 T 细胞不表达。CTLA-4 与 CD28 分子有一定的同源性,其胞外区结构域也能与 APC 表面 B7-1/B7-2 共 刺激分子互补结合,且二者间亲和力显著高于 CD28 与 B7 分子间的亲和力;其胞 内区含免疫受体酪氨酸抑制基序/模体(immunoreceptor tyrosine-based inhibitory motif, ITIM),可传递活化抑制信号。ITIM 是免疫细胞抑制性受体 分子胞内区所携带的结构,其内所含酪氨酸残基发生磷酸化后可招募含有 SH2 结 构域的蛋白磷酸激酶,并通过脱磷酸化作用抑制信号转导。活化 T 细胞通过表面 CTLA-4 与 APC 表面相应配体 B7-1/2 (CD80/CD86)结合相互作用,使其胞内 ITIM 磷酸化后可产生活化抑制信号,阻止 T 细胞过度增殖。CD28 和 CTLA-4 在 T 细胞 活化的不同阶段发挥正负调节作用。
(3)CD40 配体(CD40 Ligand,CD40L):是表达于活化 CD4+T 细胞和部分活化 CD8+T 细胞表面的共刺激分子,为 TNF 超家族成员。活化 T 细胞表面以三聚体形式 存在的 CD40L 与 B 细胞表面 CD40 分子结合相互作用,可诱导 B 细胞产生共刺激 信号(B 细胞活化第二信号)导致 B 细胞活化、增殖分化和发生 Ig 类别转换; 活化 T 细胞表面 CD40L 与树突状细胞和巨噬细胞等 APC 表面 CD40 分子结合相互 作用,可诱导上述专职 APC 活化,使其表面 B7 分子表达上调,同时合成分泌 IL- 12 等细胞因子,有效促进 T 细胞增殖分化。
(4)诱导性共刺激分子(inducible costimulator, ICOS):主要表达于活化 T 细胞表面,为 CD28 家族成员。当活化 T 细胞通过表面 ICOS 与 APC 表面相应配 体 ICOSL(人 B7-H2、鼠 B7RP-1)结合后,可诱导活化 T 细胞合成分泌细胞因子和 促进 T 细胞增殖分化。
(5)程序性死亡(蛋白)-1(programmed death-1,PD-1):主要表达于活化 T 细胞表面,为 CD28 家族成员;PD-L1 和 PD-L2 是 PD-1 识别结合的配体,主要 表达于树突状细胞和巨噬细胞表面。活化 T 细胞通过表面 PD-1 与相应配体(PD- L1 或 PD-L2)结合相互作用,可产生活化抑制信号,阻止 T 细胞增殖分化和细胞 因子的合成分泌。
(6)淋巴细胞功能相关抗原 2(lymphocyte function associated antigen- 2,LFA-2):为单链糖蛋白,是 Ig 超家族成员;主要表达于成熟 T 细胞和部分 NK 细胞表面。LFA-2(CD2)是 T 细胞表面的黏附分子,能与 APC 表面相应黏附分子 LFA-3 结合相互作用,促进 T 细胞对 APC 表面抗原肽-MHC 分子复合物识别结合和 共刺激信号(T 细胞活化第二信号)的产生。
(7)淋巴细胞功能相关抗原-1(LFA-1):是由α链和β链组成的异二聚体, 为整合素家族成员;主要表达于成熟 T 细胞表面。LFA-1(CD11a/CD18)是 T 细 胞表面的粘附分子,能与 APC 表面相应黏附分子,即细胞间黏附分子-1、2 (intercellular adhesion molecules-1、2,ICAM-1、2)结合相互作用,促进 T 细胞对 APC 表面抗原肽-MHC 分子复合物的识别结合和共刺激信号(T 细胞活化 第二信号)的产生。
2.5 丝裂原受体
T 细胞表面具有植物血凝素受体(PHA-R)、刀豆蛋白 A 受 体(ConA-R)和与 B 细胞共有的美洲商陆受体(PWM-R);他们接受相应丝裂原刺激 后可发生非特异性有丝分裂,使众多 T 细胞转化为淋巴母细胞。在体外用 PHA 刺 激人外周血 T 细胞,观察其增殖分化程度可检测机体细胞免疫功能状态,此即淋 巴细胞转化试验。
2.6 细胞因子受体
T 细胞可表达多种与其活化、增殖、分化密切相关的细 胞因子受体(如 IL-2R、IL-4R、IL-6R、IL-12R、IFN-γR)和诱导活化 T 细胞凋亡 的 Fas 等受体。
三、T 淋巴细胞亚群
T 细胞是具有高度异质性的细胞群体,摘要介绍如下几种不同的分类方法: (1)根据 TCR 肽链组成情况的不同,可将 T 细胞分为 αβT 细胞和 γδT 细 胞;(2)根据 T 细胞是否接受过抗原刺激或接受抗原刺激后的分化情况,可将其 分为初始 T 细胞、效应 T 细胞和记忆 T 细胞;(3)根据 T 细胞内外标志和功能特 性,可将其分为 CD4+Th 细胞、CD8+CTL 细胞和 CD4+调节性 T 细胞。
3.1 αβT 细胞和 γδT 细胞
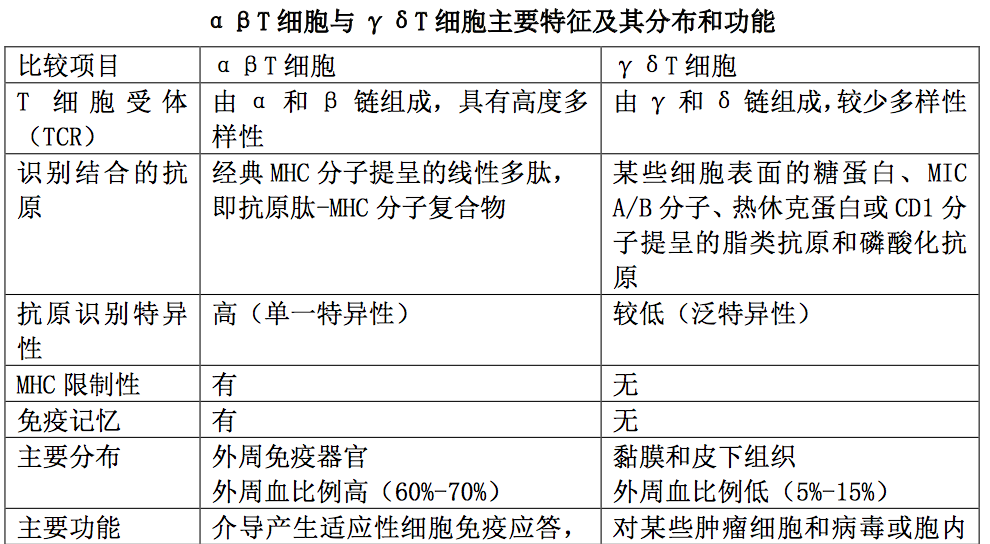
1.αβT 细胞
是执行适应性免疫应答的 T 细胞,主要分布于外周淋巴组 织和血液中;其 TCR 由 α 和 β 两条肽链组成,具有高度多样性和抗原识别特异 性。αβT 细胞只能识别 MHC 分子提呈的抗原肽,即表达在 APC 表面的抗原肽- MHC 分子复合物,并且具有自身 MHC 限制性;其主要功能是执行适应性细胞免疫 应答,辅助 B 细胞产生适应性体液免疫应答和参与免疫应答的调节。
2.γδT 细胞
是执行固有免疫应答的 T 细胞,主要分布于粘膜和皮下组 织,其 TCR 由 γ 和 δ 两条肽链组成,缺乏多样性。γδT 细胞可直接识别某些 细胞表面的糖蛋白、MIC A/B 分子、热休克蛋白或 CD1 分子提呈的脂类抗原和磷 酸化抗原,且不受 MHC 限制。他们对某些肿瘤细胞和病毒或胞内寄生菌感染的靶 细胞具有泛特异性杀伤作用,也可通过分泌不同类型的细胞因子发挥免疫调节和 介导炎症反应。αβT 细胞与 γδT 细胞主要特征及其分布和功能如表所示。

3.2 初始 T 细胞、效应 T 细胞和记忆 T 细胞
1.初始 T 细胞(naive T cell,Tn)
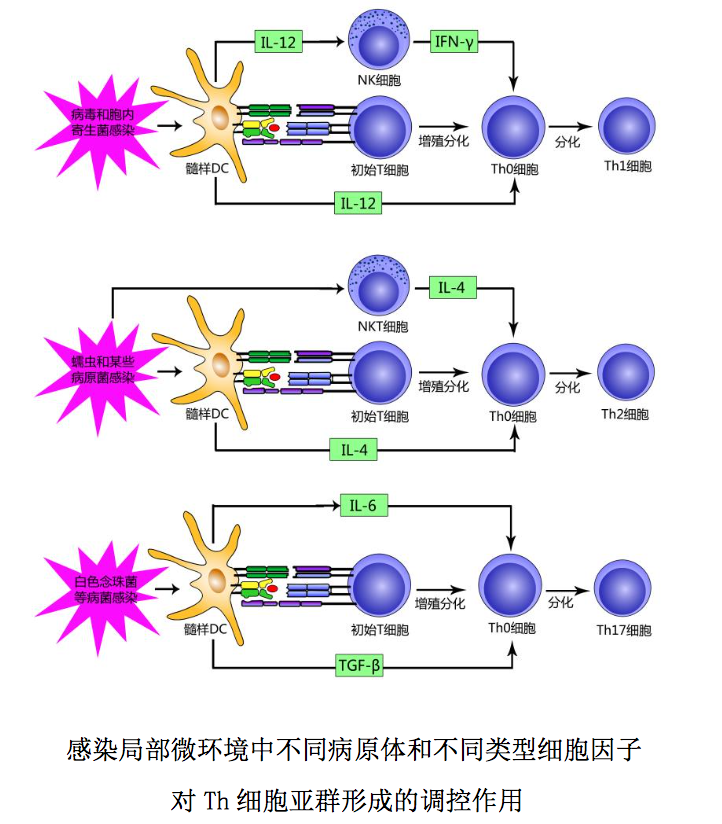
是指离开胸腺进入外周后,从未与 其特异性抗原相遇,即对相应抗原尚未产生应答的成熟 T 细胞。该种 T 细胞表 达 CD45RA 和高水平与其归巢相关的 L-选择素(CD62L)及其他粘附分子,参与 淋巴细胞再循环。在外周免疫器官,CD4+初始 T 细胞通过表面 TCR-CD3 复合体和 粘附分子与树突状细胞表面相应抗原肽-MHC 分子复合物和相应粘附分子结合相 互作用后,首先分化为 CD4+Th0 细胞;后者在局部微环境中不同病原体和/或细 胞因子作用下,可增殖分化为功能不同的 T 细胞亚群,参与适应性免疫应答,并 最终分化为效应 T 细胞和记忆 T 细胞。
2.效应 T 细胞(effector T cell,Te)
是指接受抗原刺激后,经克隆扩 增和分化形成的能够发挥免疫效应的终末 T 细胞。效应 T 细胞不表达 CD45RA 和 L-选择素(CD62L),而表达 CD45RO 和高水平 IL-2R,借此能与初始 T 细胞相区别。 效应 T 细胞能向外周炎症部位或某些器官组织迁移,而不参与淋巴细胞再循环。 CD4+/ CD8+效应 T 细胞与 APC 或肿瘤和病毒感染靶细胞表面相应抗原肽-MHC 分子 复合物特异性结合后,可通过释放 IL-2、IFN-γ、TNF-β 等多种细胞因子或分 泌穿孔素、颗粒酶等细胞毒性物质,介导产生细胞免疫效应或细胞毒作用。
3.记忆 T 细胞(memory T cell,Tm)
是指接受抗原刺激后,在增殖分化 过程中停止分化,成为静息状态的具有免疫记忆功能的长寿 T 细胞。记忆 T 细胞 介导再次免疫应答,他们对相应抗原和细胞因子的刺激较初始 T 细胞更为敏感; 在与相应抗原相遇后可迅速活化、增殖分化为效应 T 细胞和产生新的记忆 T 细 胞。记忆 T 细胞主要存在于血液和外周淋巴器官,也能向外周炎症组织等部位迁 徙;其中参与淋巴细胞再循环的记忆 T 细胞称为中央型记忆细胞(central memory cell,Tcm)。此型记忆 T 细胞为 CD45RO+ CD62L+ CCL7+ T 细胞, 能与表型 为 CD45RO- CD45RA+ CD62L+的初始 T 细胞和表型为 CD45RO+ CD62L- CCL7- 的效应 T 细胞相区别。
3.3 CD4+Th 细胞、CD8+CTL 细胞和 CD4+调节性 T 细胞
3.3.1 CD4+Th 细胞亚群及其主要功能和相互间的调控作用
CD4+Th 细胞是组成 性表达 TCRαβ 和 CD4 分子的辅助性 T 细胞(T helper cell,Th),包括 CD4+ Th1、CD4+ Th2、CD4+ Th3 、CD4+ Th17 和 CD4+ Tfh 细胞等亚群。CD4+Th 细胞亚 群形成及其产生的细胞因子和主要功能简述如下: CD4+ 初始 T 细胞接受抗原刺 激后首先增殖分化为 CD4+Th0 细胞;CD4+Th0 细胞是 CD4+ Th 细胞亚群共同的前体 细胞,可表达多种不同类型的细胞因子受体;在局部微环境中相关细胞因子作用 下,CD4+Th0 细胞可分化为如下几种 Th 细胞亚群。
(1)CD4+ Th1 细胞:CD4+Th0 细胞在 IL-12 和 IFN-γ 作用下,可分化为转 录因子 T-bet+的 CD4+Th1 细胞。该种 Th 细胞主要分泌 IFN-γ、IL-2 和 TNF-β 等 Th1 型细胞因子,参与细胞免疫应答,具有抗胞内病原体感染的免疫作用;也 参与某些自身免疫性疾病,如类风湿性关节炎(RA)的发生发展和病理损伤。
(2)CD4+ Th2 细胞:CD4+Th0 细胞在 IL-4 作用下,可分化为转录因子 Gata3+ 的 CD4+Th2 细胞。该种 Th 细胞主要分泌 IL-4、IL-5、IL-10 和 TGF-β 等 Th2 型 细胞因子,参与体液免疫应答,可诱导 B 细胞增殖分化产生抗体,具有抗胞外病 原体感染的免疫作用;也参与哮喘等变态反应性疾病的发生发展和病理损伤。
(3)CD4+ Th3 细胞:CD4+Th0 细胞在 TGF-β 作用下,可分化为转录因子 FoxP3+的 CD4+Th3 细胞(为诱导性调节 T 细胞),主要分泌 TGF-β,具有免疫负 调节作用,可抑制细胞和体液免疫应答。Th3 细胞功能过高或过低与肿瘤或自身 免疫性疾病的发生发展相关。
(4)CD4+ Th17 细胞:CD4+Th0 细胞在 TGF-β、IL-6 和 IL-23 作用下,可分 化为转录因子 RORγt+的 CD4+Th17 细胞。该种 Th 细胞主要分泌 IL-17、IL-21 和 IL-22 等促炎细胞因子,具有抗真菌和抗胞外细菌感染的免疫作用;也参与某些 炎症性疾病,如炎症性肠炎、银屑病的发生发展和病理损伤。
(5)CD4+ Tfh 细胞:是新近发现的一种称之为滤泡辅助性 T 细胞(T Folleicular helper cell,Tfh)的 CD4+ Th 细胞亚群。CD4+Th0 细胞在 IL-6 和 IL-21 作用下,可分化为转录因子 BcL-6+的 CD4+ Tfh 细胞。该种 Th 细胞高表达 趋化因子受体 CXCR5 及共刺激分子 CD40 和 ICOS;当他们迁移到淋巴滤泡后,能 与 B 细胞表面 CD40L 和 ICOSL 结合相互作用有效激活 B 细胞,并通过合成分泌、 IL-21 、IL-10 和 IL-4 等细胞因子,促进 B 细胞增殖分化产生抗体和发生 Ig 类别转换。Tfh 细胞功能过高可诱导产生大量自身抗体,引发系统性红斑狼疮(SLE) 等自身免疫性疾病;功能过低则可引发抗体缺失或低下为特征的免疫缺陷病。

CD4+初始 T 细胞的分化方向与其所在局部微环境中病原体种类、固有免疫细 胞及其分泌的细胞因子有关,举例简述如下: 1 髓样DC接受结核杆菌等胞内 寄生菌或某些病毒刺激后产生的以 IL-12 为主的细胞因子,可使 NK 细胞活化释 放 IFN-γ,并能与 IFN-γ 协同作用诱导 CD4+Th0 细胞向 CD4+Th1 细胞分化。2 髓样 DC、NKT 细胞和肥大细胞接受蠕虫等胞外病原体刺激后产生的以 IL-4 为主 的细胞因子,可诱导CD4+Th0细胞向CD4+Th2细胞分化。3 髓样DC接受白色念 珠菌等病原菌刺激后产生的 IL-6 和 TGF-β 等细胞因子,可诱导 CD4+Th0 细胞向 CD4+Th17 细胞分化。
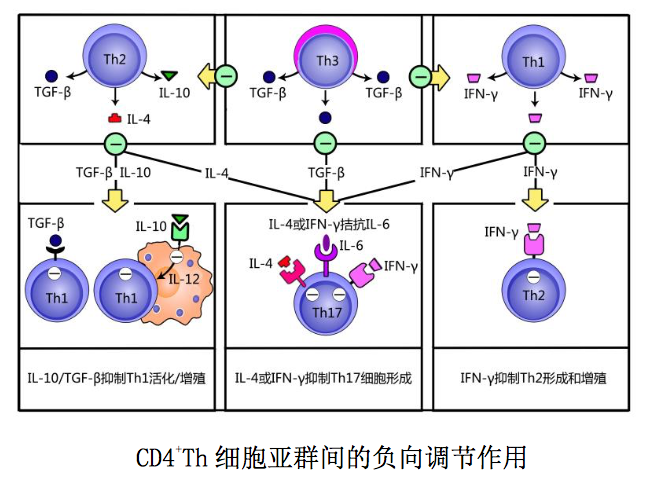
某些 CD4+T 细胞可通过产生不同类型的细胞因子对其他 CD4+T 细胞的形成或 增殖产生负向调节作用(图 10-8),举例简述如下:○1 在非感染自稳状态下, CD4+Th3 细胞和其他 Treg 细胞可通过分泌 TGF-β 抑制 CD4+初始 T 细胞活化的作 用方式,影响 Th1/Th2/Th17 等 CD4+T 细胞亚群的形成。2在感染情况下,髓样 DC 分泌的 IL-6 是诱导初始 T 细胞分化为 Th17 细胞的关键性细胞因子;活化 Th1 或 Th2 细胞产生的 IFN-γ 或 IL-4 可通过对 IL-6 的拮抗作用,抑制 Th17 细胞 的形成。3活化 Th1 细胞产生的 IFN-γ 可抑制 Th2 细胞增殖。4活化 Th2 细胞 通过分泌 IL-10 抑制巨噬细胞产生 IL-12,阻抑 Th1 细胞的形成;通过分泌 TGF- β 抑制 Th1 细胞增殖。
- 细胞毒性 T 细胞(cytotoxic T lymphocyte,CTL 或 Tc) 是组成性表 达 TCRαβ和 CD8 分子的 T 细胞;其表面 TCR-CD3 复合体识别抗原受 MHCI类分 子限制,即只能识别结合 APC 或靶细胞表面 MHCI类分子提呈的抗原肽。CD8+CTL 的主要作用是特异性杀伤某些肿瘤和病毒感染的靶细胞,他们通过表面 TCR-CD3 复合体与靶细胞表面相应抗原肽-MHCI类分子复合物特异性结合后,可通过如下 与 NK 细胞相同的作用机制产生细胞毒作用:(1)脱颗粒释放穿孔素和颗粒酶, 使靶细胞溶解破坏或发生凋亡;(2)高表达 FasL 和分泌大量 TNF-α 诱导靶细胞 凋亡。在杀伤靶细胞过程中,CD8+CTL 与 NK 细胞类似也能连续杀伤多个靶细胞。 此外, CD8+CTL还可通过分泌IL-2、IFN-γ、TNF-β等Th1型细胞因子或IL-4、 5、10 和 IL-13 等 Th2 型细胞因子发挥免疫调节作用。
3.3.2 细胞毒性 T 细胞(cytotoxic T lymphocyte,CTL 或 Tc)
是组成性表 达 TCRαβ和 CD8 分子的 T 细胞;其表面 TCR-CD3 复合体识别抗原受 MHCI类分 子限制,即只能识别结合 APC 或靶细胞表面 MHCI类分子提呈的抗原肽。CD8+CTL 的主要作用是特异性杀伤某些肿瘤和病毒感染的靶细胞,他们通过表面 TCR-CD3 复合体与靶细胞表面相应抗原肽-MHCI类分子复合物特异性结合后,可通过如下 与 NK 细胞相同的作用机制产生细胞毒作用:(1)脱颗粒释放穿孔素和颗粒酶, 使靶细胞溶解破坏或发生凋亡;(2)高表达 FasL 和分泌大量 TNF-α 诱导靶细胞 凋亡。在杀伤靶细胞过程中,CD8+CTL 与 NK 细胞类似也能连续杀伤多个靶细胞。 此外, CD8+CTL还可通过分泌IL-2、IFN-γ、TNF-β等Th1型细胞因子或IL-4、 5、10 和 IL-13 等 Th2 型细胞因子发挥免疫调节作用。
3.3.3 调节性 T 细胞(regulatory T cell,Treg)
是一类具有负向调节作用 的 CD4+T 细胞,包括自然调节 T 细胞和诱导性调节 T 细胞。此类 T 细胞约占外周 血 CD4+T 细胞总数的 5~10%;对抗原的识别具有特异性,但活化后对其他免疫细 胞的抑制作用是非特异性的,且不受 MHC 限制。
(1)自然调节 T 细胞(natural occurring regulatory T cell, nTreg): 是指在胸腺中分化而成,可组成性表达 CD4、CD25、CTLA-4 等膜分子和胞质转录 因子FoxP3的具有免疫抑制作用的CD4+ CD25+ FoxP3+调节T细胞。此类调节T细 胞可被自身抗原激活,主要通过与树突状细胞密切接触作用方式发挥负向调节作 用,也可通过释放 TGF-β和 IL-10 等抑制性细胞因子发挥负向免疫调节作用。 nTreg 作用机制复杂,择要简介如下:1nTreg 通过表面 TCR-CD3 复合受体与 APC 表面相应自身抗原肽-MHCII类分子复合物特异性结合后,可凭借表面高浓度抑制 性共刺激分子 CTLA-4 与自身反应性 T 细胞表面共刺激分子 CD28 竞争结合 APC 表 面相应共用配体 B7 分子的作用方式,使上述自身反应性 T 细胞因未能获得有效 共刺激信号而处于活化无能(anergy)状态。2nTreg 通过表面高浓度 CTLA-4 与 未成熟 DC 表面相应配体 B7 分子高亲和力结合,可诱导上述 DC 表达吲哚胺 2,3- 双加氧酶(indoleamine 2,3-dioxygenase,IDO),后者能够降解 T 细胞活化所必 需的色氨酸,并由此导致 T 细胞失活和发生凋亡。3nTreg 在与未成熟 DC 结合 相互作用过程中,可表达一种称为淋巴细胞活化基因-3(lymphocyte activation gene-3, LAG3)的跨膜蛋白;LAG3 跨膜蛋白(CD223)与 CD4 分子同源,能与未成 熟 DC 表面 MHCII类分子高亲和力结合,并通过干扰 ITAM 信号转导作用方式对未 成熟DC活化产生抑制作用,使其不能有效激活CD4+T细胞。4 nTreg可通过表 面膜型 TGF-β与 DC 表面相应配体密切结合抑制 DC 活化;也可通过合成分泌的 TGF-β与 DC 表面相应配体结合抑制 DC 活化,使其不能有效激活 CD4+T 细胞。
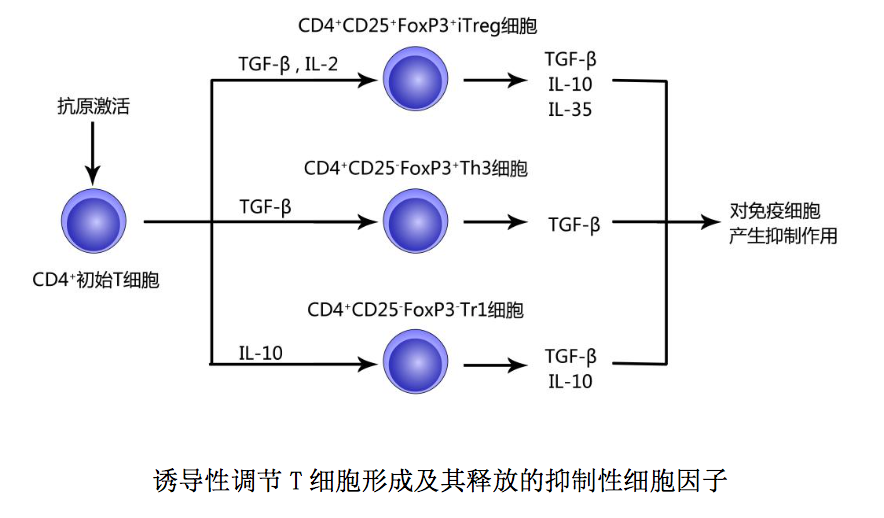
(2) 诱导性调节 T 细胞(induced regulatory T cells,iTreg ):是指外 周免疫器官和感染组织部位 CD4+初始 T 细胞接受外来抗原刺激后,在某些抑制性 细胞因子诱导下形成的对多种免疫细胞具有抑制作用的 CD4+调节 T 细胞。此类调 节性 CD4+T 细胞主要通过释放抑制性细胞因子对免疫细胞产生抑制作用。 诱导性调节 T 细胞包括以下三种:○1 CD4+ CD25+ FoxP3+诱导性调节 T 细胞 (iTreg):是抗原活化的 CD4+初始 T 细胞在 TGF-β和 IL-2 作用下形成的调节 T 细胞,主要通过分泌 TGF-β、IL-10 和 IL-35 对免疫细胞产生抑制作用;○2 CD4+ CD25- FoxP3+ Th3 细胞:是抗原活化的 CD4+初始 T 细胞在 TGF-β诱导下形成的调 节 T 细胞,主要通过分泌 TGF-β对免疫细胞产生抑制作用;○3 CD4+ CD25- FoxP3- I 型调节 T 细胞(Tr1 细胞):是抗原活化的 CD4+初始 T 细胞在 IL-10 诱导下形成 的调节 T 细胞,主要通过分泌 IL-10 和 TGF-β对免疫细胞产生抑制作用。 上述抑制性细胞因子作用简述如下:1IL-10 可通过下调 APC 表面共刺激分 子和 MHCII类分子表达作用方式,抑制 DC 成熟和抗原提呈;可通过抑制 DC 或其 他 APC 释放促炎细胞因子,下调或抑制炎症反应;可通过抑制 T 细胞和单核细胞 合成分泌细胞因子,使 CD4+/CD8+T 细胞处于无能状态;可通过抑制致敏肥大细胞 脱颗粒或诱导 B 细胞发生类别转换(IgEIgG4)作用方式,下调 Th2 细胞介导 的超敏反应。2TGF-β可通过与 APC 表面相应配体结合抑制 DC 活化和成熟;可 通过对 IL-2 产生的干扰作用抑制 T 细胞增殖;可通过对 T-bet/STAT4 或 GATA3/NFAT 功能的干扰作用抑制初始 T 细胞向 Th1 或 Th2 细胞分化,下调适应 性免疫应答。
参考资料
- 《医学免疫学》课件 新乡医学院 宋向凤、张国俊、徐春阳、孙爱平、孙书明、赵铁锁等老师
