【1.5】氨基酸温度因子 b-factor
B-Factor 值,也叫“温度因子” ,是另一个描述蛋白质灵活性的指标。如果 说 RMSF反映的是蛋白质的动态灵活性,那么 B-Factor 体现的则是蛋白质的静 态灵活性。 B-Factor 值是从蛋白质的 X 光衍射晶体结构的数据中获得的,它反 映了原子的电子层的离散程度。一个氨基酸的 B-Factor 值通过平均组成氨基酸 的所有原子的 B-Factor 来获得。 B-FITTER就是这样一个软件,从蛋白质的三级 晶体结构中提取原子 B-Factor 数据,然后平均,获得所有残基 B-Factor 值,以 此衡量蛋白质的灵活性。 但在进行蛋白质柔性区域的预测的过程中需要注意, 即 便同一种蛋白质,从不同的晶体结构中获得的 B-Factor值是有所不同的,B-Factor 值跟蛋白质的结晶质量、结构的分辨率有很大的关系。
在PDB文件中,红色框框中标记的即为b-factor值
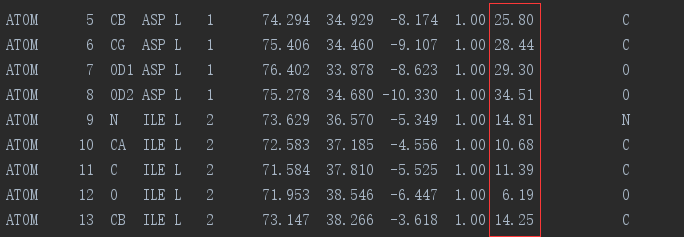
组成蛋白的氨基酸的 B-Factor 值越大,表示该氨基酸对于稳定蛋白的作用越小。因此将组成蛋白的氨基酸的 B-Factor 值进行排序,然后将 B-Factor 值高 的氨基酸替换为 B-factor 值低的氨基酸,可以提高蛋白的热稳定性。 具体的操 作方法大致分为四种,分别是:
- B-Factor 值与 Rosetta design配合确定突变位点与 突变氨基酸
- B-Factor 值与特殊结构的同源对比确定突变位点与突变氨基酸
- B-Factor 值与饱和突变
- B-Factor 值与迭代饱和突变。
二、降低氨基酸b-factor的方法
2.1 B-Factor与 Rosetta design
B-Factor 与 Rosetta design的结合使用是当前最高效的方法之一,首先计算 出组成蛋白质的所有氨基酸的 B-Factor 值,然后选出 B-Factor 值最高的氨基酸, 将其输入到 Rosetta之中,经过 Rosetta计算,确定将 B-Factor值高的氨基酸突变 为何种氨基酸,然后通过定点突变得到突变株。
怎么知道替换以后的b-factor值呢???
参考文献
1.Enhancing the Thermostability of Serratia plymuthica Sucrose Isomerase Using ·B-Factor-Directed Mutagenesis . Xuguo Duan , Sheng Cheng , Yixin , Jing Wu
将沙雷氏菌 AS9(AS9 Pal1)的蔗糖异构酶在大肠杆菌 BL21(DE3)中表达, 在 45°C下 AS9 PalI的半衰期为 20 分钟,表明其不稳定。为了改善其热稳定性, 选择具有更高 B-Factor 的 6 个氨基酸残基作为定点诱变的靶, 并且 RosettaDesign 服务器设计 6 个突变体(E175N,K576D,K174D,G176D,S575D和 N577K)。E175N 和 K576D突变体在初步实验中表现出改善的热稳定性,因此构建了双突变体 E175N / K576D。 详细表征了这三种突变体 (E175N,K576D,E175N / K576D)。 结 果表明,与野生型酶(30°C)相比,三种突变体表现出稍微升高的最适温度 ( 35°C)。 突变体也具有相同的最佳 pH 6.0,其与野生型酶的相同。 在 45°C下, E175N, K576D和 E175N / K576D突变体的半衰期分别为野生型酶的半衰期的 2.30,1.78 和 7.65 倍。动力学研究表明, E175N,K576D和 E175N / K576D突变体的 K m 值分 别下降了 6.6%, 2.0%和 11.0%,与野生型酶相比 ,其 k cat / K m 值分别增加了 38.2%,4.2%和 19.4%。
2.Development of thermostable lipase B from Candida antarctica (CalB) through in silico design employing B-factor and RosettaDesign. Hyun Sook Kim, Quang Anh Tuan Le, Yong Hwan Kim
本文采用了理性设计的方法来开发热稳定良好的 CalB酶。这种方法的第一 步是选择合适的网站显示高 B-Factor 值。基于从 X 射线数据获得的原子位移参数 选择这些位点。 然后用其它氨基酸取代所选择的位点, 所述其他氨基酸通过使用 ‘RosettaDesign的’计算建模确定一个比原始氨基酸更刚性的氨基酸。此外,包装 分析工具“ Voronoia”指出,野生型 CalB1249残基的一个空腔在 CalB R249L中消 失,其可以是 CalB R249L突变体与野生型 CalB相
2.2 B-Factor 与特殊结构的同源对比
B-Factor 与特殊结构的同源对比结合的方法,首先计算出组成蛋白质的所有 氨基酸的 B-Factor值,然后选出 B-Factor值高的氨基酸区域。然后与同源的热稳 定性好的蛋白进行同源对比。 比较出哪些氨基酸有助于提高蛋白的热稳定性。 然 后通过定点突变实现氨基酸的替换。 还有就是在蛋白的特殊区域内, 用特殊的氨 基酸替换原有氨基酸可以提高蛋白的热稳定性。
参考文献:
- Polarity Alteration of Calcium Site Induces a Hydrophobic Interaction Network and Enhances Cel9A Endoglucanase Thermostability . Hsiu-Jung Wang , Yu-Yuan Hsiao, Yu-Pei Chen , Tien-Yang Ma , Ching-Ping Tseng
钙结合位点通过稳定天然褶皱和改变局部构象来控制蛋白质的热稳定性和 活性。酸热脂环酸杆菌在热酸性条件下存活并产生内切葡聚糖酶 Cel9A ( AaCel9A),其在催化裂口附近含有钙结合位点 (Ser465-Val470)。我们发现 Ca 2+ 通过驱动构象变化诱导钙位点和其附近区域之间的疏水相互作用。在高 B-Factor 区域可以进通过将周围的极性残基替换为疏水残基而进一步增强疏水相互作用, 从而影响酶热稳定性和活性。 因此,其侧链直接连接 Ca2+的钙结合蛋白残基 Asp468以及 AaCel9A的 Asp469和 Asp471分别被丙氨酸和缬氨酸取代。 与野生 型相比, 突变体 D468V 和 D468A 解链温度分别上升 4.5°C和 5.9°C ; 并且在 75°C 下的酶半衰期增加了约 13倍。AaCel9A和 GH9家族的其他内切葡聚糖酶之间的 结构比较表明, AaCel9A钙位点区域的稳定性在高温下在 GH9 内切葡聚糖酶催 化中起重要作用。
2.Engineering the thermostability of -glucuronβidase from Penicillium purpurogenum Li-3 by loop transplant . Xudong Feng, Heng Tang, Beijia Han,Liang Zhang , Bo Lv , Chun Li
在本研究中,提出环移植策略,以提高在大肠杆菌中表达的产黄青霉 Li-3β - 葡糖醛酸糖苷酶(缩写为 PGUS-E)的热稳定性。首先,利用 B-Factor 值,确定 待替换的 PGUS-E的三个不稳定表面环:环 205-211,258-263和 25-31。然后,基 于 B-Factor分析,与 PGUS-E具有低同源性的两种典型的嗜热糖苷酶进行同源对 比,从中选择 8 个稳定的取代环 (小于 25%)。通过分析这些稳定环的共同特征, 发现它们共享共同的残基骨架 DXXT(X X)R,基于此,还手动设计了三个嵌合环: RSQTSN,D RSSTQRD和 DDQTSR。通过同源结构引入所有这些环以代替 PGUS-E 的不稳定环, 选择具有增加氢键数目的突变体和与局部突变区域的良好相容性的 突变体进一步进行实验验证。通过使用该策略,实验产生了 10 种突变体,其中 获得了在 70°C下比野生型( 8.5 分钟)高 11.8,3.3 和 9.4 倍半衰期的苏氨酸 M1 , M3 和 M8。最后, MD 模拟表明增加的氢键,减少 N-末端的柔性和增加的 π-π 堆积相互作用负责改善的热稳定性。
3 . 理性设计改造牛肠激酶的热稳定性 . 郭 超、王志彦、甘一如、李 丹、邓 勇、于浩然、黄 鹤
以牛肠激酶作为研究对象, 利用理性设计的方法提高其热稳定性。 首先通过 分子动力学模拟软件 Gromacs v 4. 5.5,FlexService以及 B-FITTER软件预测出了 肠激酶的柔性区 Fragment 64~69,Fragment 85 ~90;然后结合 β-转角序列统 计学信息以及引入位置原有的残基不参与形成氢键的原则, 确立了 3 个突变位点 S67P,R87P 以及 Y136P;通过 Quik Change TM 定点突变的方法引入突变位点, 并进行了酶热稳定性分析 . β -转角是最小的蛋白二级结构,由于其不存在复杂的相互作用力,通常处 于蛋白柔性较高的 loop 内。Watanabe 等研究表明,在 β-转角或无规卷曲位 置引入脯氨酸可以降低蛋白质去折叠时的骨架熵提高蛋白质的热稳定性。
2.3 B-Factor 与饱和突变
B-Factor 与饱和突变结合的方法也是比较普遍的方法之一,首先计算出组成 蛋白质的所有氨基酸的 B-Factor 值,然后选出 B-Factor值高的氨基酸区域。然后 利用饱和突变, 构建每一个高 B-Factor值氨基酸位点的突变体文库。 然后利用适 合的筛选方法筛选出理想的突变菌株。
1.High-throughput screening of B factor saturation mutated Rhizomucor miehei Lipase thermostability based on synthetic reaction . Jun-hui Zhang , Ying Lin , Yu-fei Sun , Yan-rui Ye , Sui-ping Zheng , Shuang-yan Han
酶的定向进化需要高通量筛选方法从具有数千个突变体的文库中发现阳性 突变体。本文提供在菌落板上的筛选培养基中使用 pH 指示剂的方法,其可以容 易地以高通量方式筛选脂肪酶合成活性, 而不需要昂贵的仪器或特殊试剂。 通过 在加入合成底物之前加热菌落平板来评价脂肪酶热稳定性。 脂肪酶残留合成反应 通过集落附近的晕轮显现。 因此,突变体文库可以从数千个转化体中减少到几十 个。进一步确认后, 我们可以生产在合成反应中具有增强的热稳定性的重组脂肪 酶菌株。基于这种新方法, 我们成功地从根据 R.miehei 脂肪酶氨基酸残基 B-Factor 的饱和突变文库中筛选具有较高合成活性和热稳定性的脂肪酶, 在培养皿水平的 pH 指示剂筛选方法中,获得了在蛋白质柔性和刚性之间实现平衡的双突变体, 并且在合成反应中显示更高的热稳定性。
2.Enhancing RGI lyase thermostability by targeted single point mutations .In ê s R. Silva , Dorte M. Larsen, Carsten Jers ,Patrick Derkx ,Anne S. Meyer , J?rn D.Mikkelsen
本文首先通过组合几种方法,即同源对比方法, PoPMuSiC可(以预测蛋白质 中所有可能的单突变的折叠自由能( ddG)的变化 )和 B-Factor建模,通过创建用 于位点饱和诱变的文库来鉴定用于该置换的最佳的氨基酸。 这种方法基于这些原 则,即这些不同方法的结果的组合将优化取代最小数目。 该方法包括以下: 预测 令人满意的替代的氨基酸, 构建每个的位点饱和的突变体文库, 并且筛选评估获 得的每个酶突变体产物的活性和热稳定性。
3.Improving the thermostability of lipase Lip2 from Yarrowia lipolytica Sai Wen , Tianwei Tan , Huimin Zhao
解脂耶氏酵母脂肪酶 Lip2( Y1Lip2)是高度通用的生物催化剂。然而其实际 使用经常受到其低稳定性的妨碍。 本文使用三种互补蛋白质工程策略来改善该酶 的热稳定性。 第一种是易错 PCR定向进化, 第二个策略是使用 B-Factor迭代测试 ( B-FIT)的半理性设计,最终发现两个热稳定的 YlLip2 变体,其显示在 50°C下 热失活的半衰期比野生型酶分别长 2 倍和 5 倍。第三种策略是使用定点诱变来组合前两种策略中鉴定的所有热稳定突变,与野生型酶相比,将 Y1Lip2的热失活 半衰期提高 7倍。
在本研究中,我们使用基于易错 PCR的定向进化和 B-FIT策略成功识别有助 于 YlLip2的结构稳定性的三个热稳定点突变。 这些热稳定化突变的进一步组合 产生了与野生型脂肪酶相比具有 7 倍增加的热失活半衰期的热稳定性 Y1Lip2变 体。
2.4 B-Factor 与迭代饱和突变
B-Factor 与迭代饱和突变结合的方法是有效方法之一,首先计算出组成蛋白质的所有氨基酸的 B-Factor 值,然后选出 B-Factor 值高的氨基酸区域。然后选择 包含一个或多个氨基酸的这些位点用于饱和诱变, 形成小聚焦文库。 突变效果最佳的突变体随后用作进行第二轮饱和诱变的模板, 通过重复地选择最佳的突变体 来选择理想的突变体
参考文献:
1.Iterative Saturation Mutagenesis on the Basis of B Factors as a Strategy for Increasing Protein Thermostability . Manfred T. Reetz, Jos D. Carballeira, and Andreas Vogel
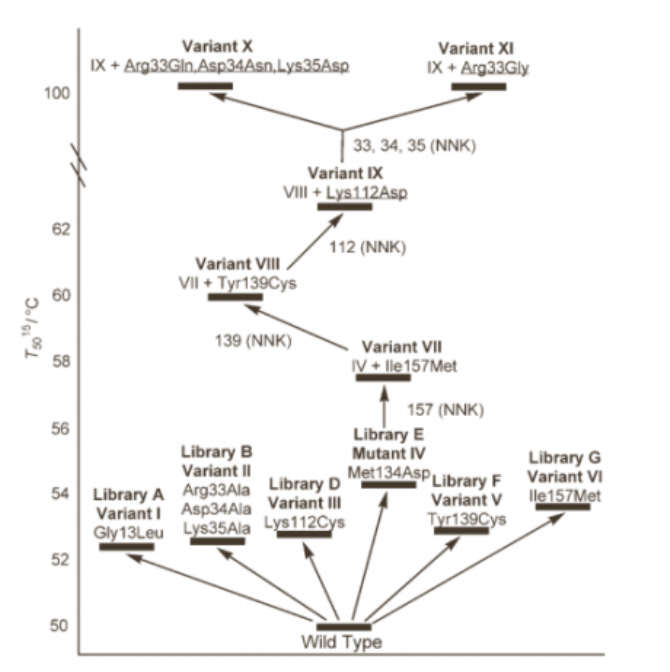
本文使用一种新的方法来选择氨基酸交换的位点, 加上有效的进化策略, 从 而消除了筛选大型文库的需要但显着增加了热稳定。通过增加适当部位的刚性, 增强嗜温酶的热稳定性是可能的。 关键是以有效的方式实现热稳定性的提高, 而 不损失酶活。首先,基于从 X 射线数据获得的原子位移参数,即 B-Factor,选择 要增加刚度的合适位置。 它们反映了相对于它们的平衡位置的作为无序热运动的 结果的原子电子密度。 因此,我们只针对显示最高 B-Factor的蛋白质中的那些氨 基酸, 对应于最显着的热运动程度, 因此具有灵活性。 然后选择包含一个或多个 氨基酸的这些位点用于饱和诱变, 形成小聚焦文库。 最佳命中的基因随后用作在 其它选择的位点之一进行第二轮饱和诱变的模板, 通过重复地聚焦于其余站点来继续(一个站点可以被不止一次地考虑) 。这种收敛过程通过以迭代方式组合敏 感的氨基酸位点(非特异性突变)来最大化添加和 / 或协作效应的可能性。这种 “迭代饱和诱变”的概念是成功的关键。 它是一种新的进化策略,也形成了迭 代 CASTing(CAST=组合活性位点饱和试验)作为增加酶的对映选择性的手段的 基础。在这种情况下应用的迭代饱和诱变提供了变体, 所述变体统计上不容易通 过先前用于产生热稳定性的传统随机诱变方法获得。
2.Enhanced Enzyme Kinetic Stability by Increasing Rigiditywithin the Active Site Yuan Xie , Jiao An , Guangyu Yang , Geng Wu , Yong Zhang, Li Cui , and Yan Feng
在这项研究中,我们实现了南极假丝酵母脂肪酶 B( CalB)动力学稳定性的 增强,通过在活性位点突变结构柔性残基。 选择具有高 B-factor 的催化性 Ser 105 残基的六个残基进行迭代饱和诱变。 在筛选 2200 个菌落后, 我们获得了 D223G / L278M 突变体在 48°C下的半衰期增加了 13 倍和 T 50 提了高 12°C,并且在 15 分钟的热处理后酶活性降低至 50%。进一步的表征显示,酶整体对热和化学变 性的抗性也得到了改善。分析野生型 CalB和 D223G / L278M突变体的晶体结构 显示后者形成额外的主链氢键网络,其中在柔性 α10 螺旋内具有七个结构上偶 联的残基,其主要参与形成活性位点。进一步研究相对 B-factor 分布和分子动 力学模拟证实,刚性的增强降低了高温下活性位点残基的波动。
3.Biophysical characterization of mutants of Bacillus subtilis lipase evolved for thermostability: Factors contributing toincreased activity retention Wojciech Augustyniak, Agnieszka A. Brzezinska, Tjaard Pijning, Hans Wienk, Rolf Boelens, Bauke W. Dijkstra, and Manfred T. Reetz
本文提出了通过在酶中特定氨基酸位置应用的迭代饱和诱变( ISM)来产 生酶热稳定性的不同方法。 为了指导关于待改变的氨基酸位点的决定, 开发了 B-FIT方法,其中靶酶的晶体结构中具有高 B 因子的残基被认为是适当的突变 候选。这种方法的基本原理是热稳定性蛋白质比其在环境温度下的嗜温相应物 更高的刚性, 并且认为在高 B-Factor 所指示的柔性区域的靶向诱变可导致硬化 并因此导致热稳定化。 作为在一个 X 射线结构中显示最高 B-Factor 的位点的几 轮 ISM的结果,我们获得了在高于 65°C的温度下处理后保留显着活性的突变 体,而野生型( WT)脂肪酶丧失所有活性。
总结
利用 B-Factor 值来预测蛋白质的柔性区域, 进而突变增强蛋白质的热稳定 性,主要的操作方法大致分为四种,分别是 B-Factor 值与 Rosetta design配合 确定突变位点与突变氨基酸, B-Factor 值与特殊结构的同源对比确定突变位点 与突变氨基酸, B-Factor 值与饱和突变, B-Factor 值与迭代饱和突变。其中 B-Factor 值与 Rosetta design配合的方法是最高效节省的方法, B-Factor 值与 迭代饱和突变的配合是提升效果比较高的一种方法。
参考资料
