【3.7.5】瞬态蛋白质-蛋白质复合物的界面残基在蛋白质间和蛋白质内相互作用
sam点评:
正如题目提到的,该文的亮点在于:通过数据分析发现,瞬时蛋白复合物中蛋白质间interface的残基,也在蛋白质内相互作用起到重要作用。 (第一次读这个题目居然有点绕。。)
背景 :蛋白质-蛋白质相互作用对于正常的生物学过程和调节影响基因表达和功能的细胞反应至关重要。先前的一些研究强调了蛋白质-蛋白质复合物界面上的残基在赋予复合物稳定性和特异性方面的作用。蛋白质中的界面残基因其与侧链和主链原子与相互作用的蛋白质的相互作用而众所周知。但是,尚不清楚蛋白质间相互作用中涉及界面残基的蛋白质间相互作用的程度及其相对于蛋白质间相互作用的相对贡献。本文使用已知的3-D结构的蛋白质-蛋白质复合物数据集探讨了此功能。
结果: 我们已经分析了45种瞬时蛋白质-蛋白质复合物结构的数据集,其中至少一种相互作用的蛋白质具有已知结构,也可以非结合形式使用。我们观察到,大部分界面残基(2137个界面残基中的1608个残基,占75%)同时参与了蛋白质内部和蛋白质间的相互作用。发现这种涉及分叉相互作用的界面残基的氨基酸倾向与发生在蛋白质-蛋白质界面的一般倾向高度相似。最后,我们观察到也以非复合形式观察到界面残基的大部分蛋白内相互作用(分叉相互作用)(83%)。
结论: 据我们所知,这是首次显示,除蛋白质间相互作用外,绝大多数蛋白质-蛋白质界面残基均参与广泛的蛋白质内相互作用。对于大多数这样的界面残基,在瞬态相互作用期间,三级结构中的微环境是预先形成的,并在与其同源伴侣形成复合物后保留。
一、背景
两种或多种蛋白质之间的缔合(Association)是许多细胞过程的核心[1]。这些关联在蛋白质之间的结合和关联形式的蛋白质的三维(3-D)取向方面都是高度特异性的[2]。此外,这些复合物中的许多在本质上是瞬时的。即,关联和解除关联( association and disassociation )是连续的过程。
在瞬时蛋白质-蛋白质复合物中,蛋白质之间的缔合强度也是一个重要特征,必须根据所涉及的蛋白质和蛋白质之间的缔合前所需的缔合时间,将其维持在精确的水平上[3]。实际上,一些有助于该复合物稳定的重要能量的界面残基被称为“热点”。已知此类残基的突变会影响所涉及蛋白质之间的结合亲和力[4,5,6]。
显然,蛋白质-蛋白质界面上的残基在赋予复合物正确的稳定性水平以及赋予蛋白质与其精确的3D结构之间的关联特异性方面起着至关重要的作用。此类残基的突变会损害相关蛋白质及其复合物的稳定性和/或特异性,从而导致疾病状态[7]和其他性质发生变化[8]。当蛋白质处于解离形式时,瞬态蛋白质-蛋白质复合物中的界面残基起什么作用?详细的分析表明,流动性有限的界面残基的一个子集可充当锚,从而有助于蛋白质之间缔合的特异性[9]。
但是,目前尚不清楚,界面残基在蛋白质内相互作用中的作用是什么?例如,诸如精氨酸的界面残基可以在蛋白质内以及与相关的蛋白质同时形成氢键。
本工作分析了蛋白质-蛋白质复合物的已知3-D结构,以了解蛋白质内界面残基的相互作用程度(蛋白质内相互作用),以及与相关蛋白质界面中的残基相互作用(蛋白质间相互作用)的程度。
二、方法
2.1 数据集
根据较早的文献,已知至少有176种瞬时蛋白质-蛋白质复合物具有结合和未结合两种形式的可用结构信息[9,10]。对于当前的分析,我们提出了一个条件:
- 即该复合物的结构应具有高分辨率(等于或优于2Å),
- 每个复合物中的两种蛋白质中至少有一种应具有游离形式的结构。
这样就得到了45条蛋白质数据库(PDB)条目,涉及114条蛋白质链,具有已知结构的蛋白质-蛋白质复合物,其中复合物中的至少一种蛋白质结构也可以非结合形式获得。我们使用这种条件是因为我们想探索结合和游离形式的涉及界面残基的蛋白内相互作用的保留程度。由于45个PDB条目中的某些对应于晶体不对称单元中一个蛋白质-蛋白质复合物的一个以上副本,因此数据集中的链数是PDB条目数的两倍以上。在这种情况下,在不对称单元中的复合物的拷贝之间发现了结构特征的一些差异。因此,当前分析中已使用所有114条链。
2.2 残基-残基相互作用的鉴定和分类
相互作用中的一对原子(分别来自蛋白质-蛋白质复合物中两种蛋白质)被认为是界面原子。使用PIC服务器[12]识别了相互作用的原子(见下文)。如果配合物中的界面原子来自残基的侧链,则该残基被认为是界面残基。复合物蛋白质中的界面残基的集合被认为是该蛋白质的界面区域(interfacial region)。
蛋白质间和蛋白质内的相互作用都使用PIC服务器识别[12]。如果相互作用涉及残基的至少一个侧链原子,则认为该残基参与相互作用,并且包括该相互作用用于进一步分析。与结合的蛋白质的侧链或主链相互作用的蛋白质中的所有侧链被认为是蛋白质-蛋白质界面残基。区分相互作用并根据结合蛋白上的类型(例如范德华,氢键,芳香族-芳香族和盐桥)进行分类。如果同时发生范德华力和氢键相互作用等,则同一残基对可能会以一种以上的形式列出。在这种情况下,具有相互作用残基的对仅计算一次,尽管同样的两个残基可能不止一个。同一残基也可能与一个或多个亚基内的一个以上残基相互作用。这样的残基-残基相互作用被分别计数。
2.3 氨基酸倾向计算
根据标准Chou-Fasman [13]类型的倾向性测量方法,测量了存在于界面上并参与分叉相互作用(蛋白质内和蛋白质间)的氨基酸的倾向性。可以通过PIC服务器的结果,通过两次单独的运行来推断这两种相互作用(一个带有“蛋白-蛋白相互作用”选项,另一个带有“蛋白内相互作用”选项)。
2.4 相互作用能的计算
PPCheck用于鉴定和定量蛋白质-蛋白质界面中的相互作用[14]。考虑到Cα-Cα距离在10Å以内的残基,并考虑了相互作用的性质来计算能量。能量以范德华力,静电力和氢键相互作用的总和来衡量。这些类型的相互作用的能量贡献是根据焓计算得出的。另外,已经使用了距离相关的电介质,并且在固定氢原子之后分析了氢键。
为了测量分叉相互作用中涉及的界面残基所具有的相互作用的强度,最初使用PIC服务器中的“蛋白内相互作用”选项来识别蛋白内相互作用。对于具有分叉相互作用的每个界面残基,单独计算所有相互作用的蛋白内残基的微环境,以进行蛋白内相互作用的PPCheck计算。在两个残基之间观察到多重相互作用的情况下,残基之间所有相互作用的总能量与残基对相关。
三、结果与讨论
3.1 蛋白质-蛋白质界面残基在蛋白质内相互作用的程度
在目前的分析中,我们使用了45种蛋白质-蛋白质复合物的数据集,这些蛋白质-蛋白质复合物的已知晶体结构(以2Å或更高分辨率确定)具有每种复合物中至少一种以非复合形式存在的蛋白质的3-D结构( 表1,附加文件1:表S1)。 我们使用未复杂的蛋白质结构来探索涉及与其他蛋白质复合形式的界面残基的蛋白质内部相互作用的保留程度。
表1. 35个 瞬时但白蛋白复合物的PDB ID
在复杂的结构中,使用PIC鉴定了蛋白质内以及结合的蛋白质之间的相互作用,例如氢键,范德华力和盐桥等[12]。分析了蛋白质-蛋白质界面上的这些残基的蛋白质内部和蛋白质间相互作用,并计算了与相互作用相关的能量。对于每个界面残基,均考虑了两类相互作用:(1)残基与结合蛋白的相互作用,以及(2)侧链与结合蛋白以及同一蛋白内的同时相互作用。第一类仅对应于跨蛋白质-蛋白质界面的相互作用。第二类是指给定侧链产生的两种相互作用。残基与同一蛋白质链中的侧链或主链原子形成的相互作用以及跨蛋白质-蛋白质界面的相互作用。因此,这两类相互作用分别称为“仅蛋白间相互作用”和“同时蛋白间和蛋白内相互作用”。
在45个复杂结构的整个数据集中的2137个界面残基中,有529个残基仅形成蛋白质间的相互作用。有趣的是,大多数1608个界面残基同时形成蛋白质间和蛋白质内相互作用(附加文件2:表S2)。因此,涉及界面残基侧链的蛋白质内相互作用也有助于蛋白质-蛋白质复合物的结构和稳定性。附加文件3中显示了涉及分叉相互作用的残基百分比分布:图S1。除五个链(对应于PDB条目2cio,1f3v,2uyz,1gl4和2fcw;附加文件2:表S2)外,大多数PDB链中的界面残基都参与了分叉的相互作用。其中,两条蛋白质链(对应于PDB代码1f3v和2fcw)与未结合的形式相比,发生了较大的构象变化(请参阅下文)。在另外两个(PDB代码1uyz和1gl4)中,另一个链主要参与分叉的交互作用。
剩余的25%的界面残基没有清晰的蛋白内相互作用,但参与蛋白间相互作用。除了Gln和Lys外,该组中涉及的残基类型列表与同时发生的内部和蛋白质相互作用(参见下面的倾向计算结果)所涉及的残基类型列表几乎相同。虽然Lys不参与蛋白内相互作用的趋势可能是由于-NH2基团在其侧链末端的局部性质,但尚不清楚为什么Gln与同时存在的蛋白内相互作用相比,显示出更高的蛋白间相互作用和蛋白质间的相互作用趋势。
3.2 残基类型同时形成蛋白内和蛋白间相互作用的可能性
在以前的几篇出版物中已经报道了在界面中出现的20种残基类型各自的可能性。在这项工作中,已经计算出20种残基类型中的每一种形成同时的蛋白质间和蛋白质内相互作用的可能性。为了保持一致性,还计算了在界面中出现的可能性,以方便进行比较(图1)。
- 除Gln和Lys以外的所有残基类型均具有大于1的倾向以形成分叉的相互作用(即同时发生的蛋白质内和蛋白质间相互作用)。因此,几乎所有残基类型都引起形成同时的蛋白内和蛋白间相互作用的趋势,而这些残基类型都显示出在蛋白-蛋白界面上发生趋势。有趣的是,这些残基类型包括疏水残基(如Leu,Phe,Trp和Met)以及极性残基(如Asp,Glu,His和Arg)。
- 还值得注意的是,尽管Gln和Lys是长侧链,但尽管它们对蛋白质间相互作用的倾向大于1,但它们对同时发生的蛋白质间和蛋白质间相互作用没有很高的偏好。相反,Glu和Arg在界面处以及同时的蛋白内和蛋白间相互作用都显示出很高的趋势。
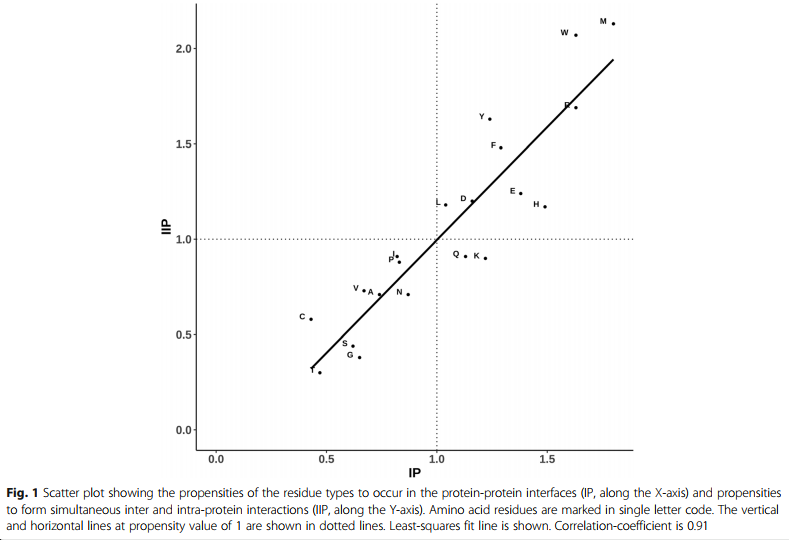
蛋白质-蛋白质界面中发生的低倾向性(<1)残基类型也显示出同时发生蛋白质内和蛋白质间相互作用的低倾向(<1)。因此,几乎不考虑残基类型,同时蛋白质内和蛋白质间相互作用的形成是界面残基的一般特征。
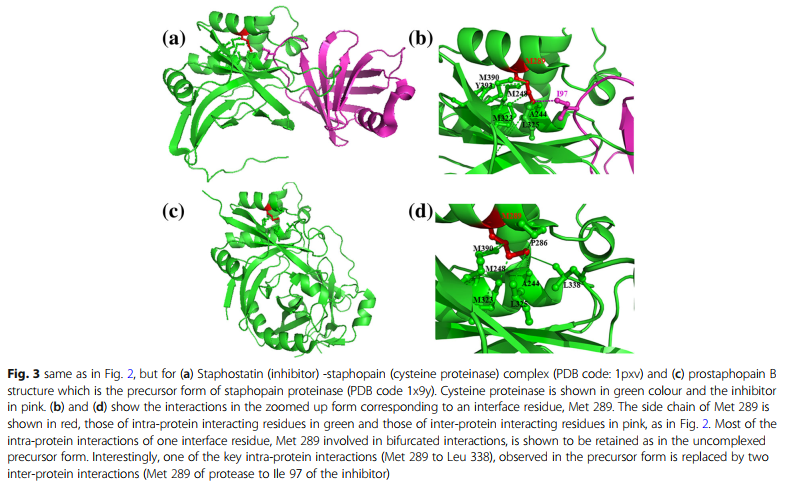
图2和图3分别显示了涉及界面残基精氨酸和蛋氨酸的同时相互作用的实例,这些相互作用分别参与了分别对应于PDB代码2es4和1pxv的蛋白质-蛋白质复合物中的分叉相互作用。如实施例所示,精氨酸与带负电荷的残基(天冬氨酸和谷氨酸)形成蛋白内和蛋白间相互作用。如第二个实例所示,蛋氨酸残基与相邻残基(蛋氨酸,脯氨酸,丙氨酸和亮氨酸/异亮氨酸)形成疏水相互作用。有趣的是,参与大多数蛋白间相互作用的残基以非复合形式参与蛋白内相互作用,反之亦然。例如,蛋白中未复合形式的Met-Leu相互作用被蛋白-蛋白复合物中的Met-Ile相互作用所取代(图3)。

3.3 残留物类型形成蛋白质内部和蛋白质间相互作用的能量贡献
使用PPCheck [14]计算与蛋白质-蛋白质复合物结构中界面残基的侧链原子与周围原子的相互作用相关的能量值。 与每个界面残基的侧链相关的相互作用的总能量表示为与蛋白间和蛋白内相互作用相关的能量之和。 图4a和b分别显示了跨整个数据集但被划分为20种残基类型中每一种的蛋白间和蛋白内相互作用能的分布。 每种残渣类型均显示了带有平均值的能量值的总范围。
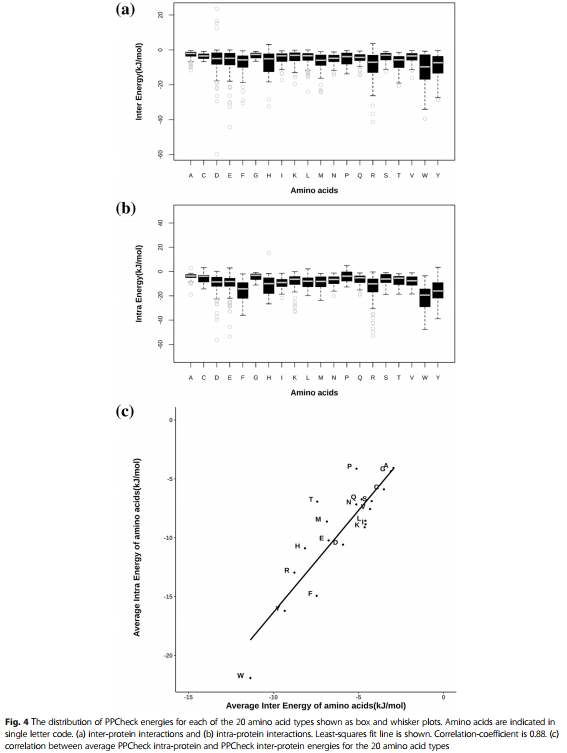
- 从图4a可以看出,与最小蛋白质间相互作用能值相关的残基类型为Trp,Tyr,His和Arg。
- 从图4b中可以看出,与最小蛋白质内相互作用能值相关的界面残基类型为Trp,Tyr,Phe,His和Arg。
图4c显示,这20种残基类型中的每一种的蛋白间和蛋白内相互作用能值均具有较好的相关性。由于大多数与最低的蛋白质内和蛋白质间相互作用能值相关的残基类型是常见的,因此可以推断出,蛋白质-蛋白质界面上优选的残基类型总体上通过蛋白质间和蛋白质间贡献了大量的稳定能量。。
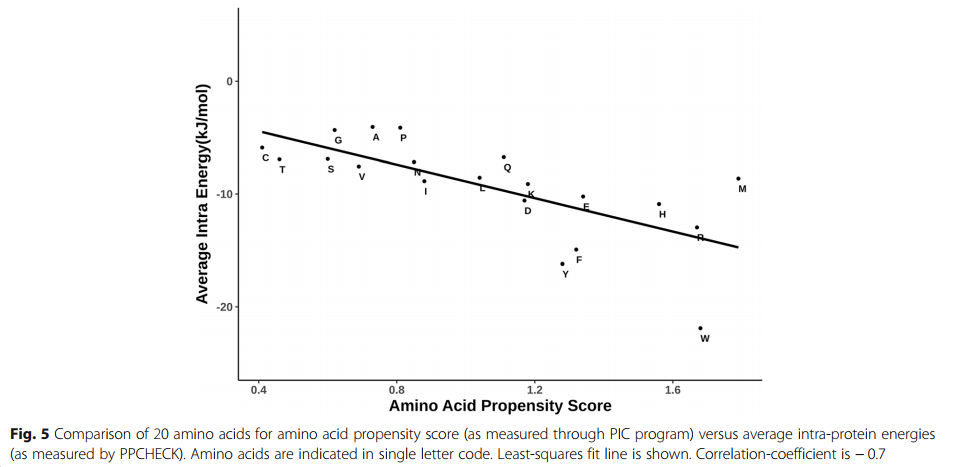
通过图5所示的散点图进一步加强了这种学习,其中沿X轴显示了界面中出现20种残基类型的倾向,沿Y轴显示了20种残基类型中每种残基的平均蛋白内相互作用能值-轴。两种分布之间的合理良好相关性证实,除蛋白质间相互作用外,蛋白质-蛋白质界面处优选的残基类型通常通过蛋白质内相互作用使复合物具有稳定性
3.4 界面残基对蛋白质内和蛋白质间相互作用能的贡献
尽管很明显,蛋白质之间的相互作用也涉及到倾向于在蛋白质-蛋白质界面上的残基类型,但尚不清楚各个界面残基对蛋白质间和蛋白质内相互作用能的贡献程度。
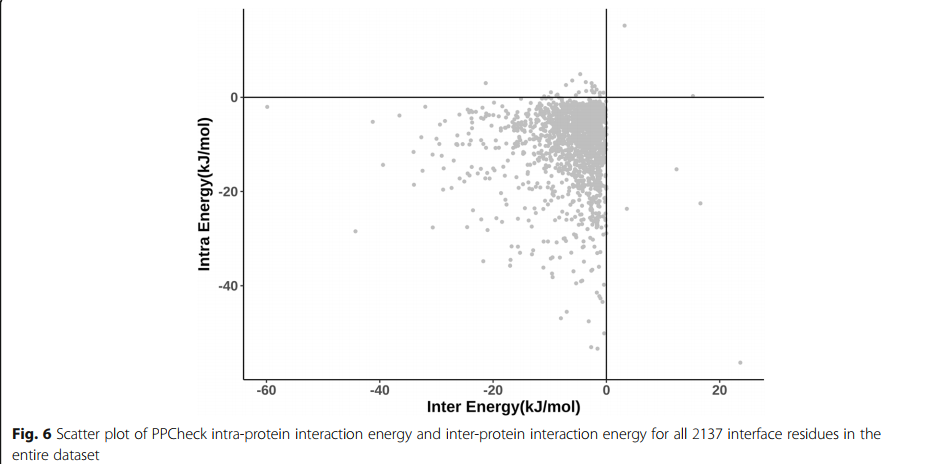
图6显示了数据集中2137个蛋白质-蛋白质界面残基的蛋白质内相互作用能和蛋白间相互作用能之间的散点图。可以注意到,对于许多残基,蛋白内和蛋白间相互作用能值非常相似。因此,数据集中的大多数界面残基贡献了蛋白质内相互作用能,几乎与它们通过蛋白质间相互作用所贡献的能量一样多。考虑到接近Y轴的点的垂直条纹,很明显,有许多残基通过蛋白质内相互作用的能量贡献远大于通过蛋白质间相互作用的能量贡献。实际上,具有相反趋势的点数,即通过蛋白间相互作用比通过蛋白内相互作用更重要的贡献显然要少得多,这表明蛋白内相互作用对蛋白-蛋白复合物的稳定性有很大贡献。
3.5 说明性例子
有趣的是,在蛋白质-蛋白质复合物结构的例子中,如图1和2所示。如图2和3所示,与蛋白质内的残基形成的相互作用主要以未结合蛋白质的形式观察到。在数据集中的大多数实例中都观察到了这种趋势。只有四种复合物显示出复合物和未复合物之间的蛋白质内相互作用保留率低(复合物fom的PDB代码1f3v,1nrj,2fcw和2vn5)。据报道这些在复杂的形成时经历巨大的结构变化。附加文件4:表S3提供了分叉相互作用中涉及的那些界面残基的结合和未结合形式的相互作用残基列表。附加文件5:表S4列出了以非复杂形式观察到的界面残基的蛋白内相互作用(参与分叉相互作用)的百分比。如果在当前分析中考虑45个瞬态复合物中的41个,除上述四种情况外,平均百分比为82.7%。这表明,即使在与伴侣蛋白复合之前,界面残基形成分叉相互作用的微环境仍保持预先形成的稳定性。这进一步表明,这种瞬时蛋白质-蛋白质复合物中的某些残基在未结合形式和结合形式之间的界面区域不会发生巨大的结构变化。这与之前的观察结果一致[9]。
四、结论
在本文中,我们证明并强调了一个事实,即蛋白质-蛋白质界面处的残基不仅通过蛋白质间相互作用,而且还通过蛋白质内相互作用对复合物的稳定性做出了重要贡献。显然,蛋白质的形状,构象,化学性质以及与蛋白质界面相关的动力学性质和程度在赋予蛋白质-蛋白质复合物稳定性和特异性方面非常重要[15]。因此,蛋白质-蛋白质界面残基的侧链起着双重作用-通过与结合蛋白的相互作用以及与蛋白质中容纳相关残基的近端原子的相互作用直接促进复合物的稳定性。
我们还显示,蛋白质内相互作用是几乎所有界面残基的普遍特征。在这种“自稳定”相互作用中,能量贡献的性质和程度在界面残基之间有所不同。蛋白内相互作用的能量贡献被证明是相当大的。具有同时发生的蛋白质内和蛋白质间相互作用的倾向的残基类型包括疏水残基Leu,Phe,Trp和Met,以及极性残基Asp,Glu,His和Arg。因此,同时的蛋白质内和蛋白质间相互作用包括各种类型,例如疏水基团和氢键之间的相互作用。
从这项工作中学到的知识鼓励人们在设计定点突变体,修补蛋白质-蛋白质复合物的稳定性/特异性和从头设计时,应考虑除蛋白质内的相互作用外,还应考虑界面残基之间的蛋白质相互作用。
参考资料
- Biology Direct volume 14, Article number: 1 (2019).Interface residues of transient protein-protein complexes have extensive intra-protein interactions apart from inter-protein interactions. https://biologydirect.biomedcentral.com/articles/10.1186/s13062-019-0232-2
