【6.1.1】CRISPR/Cas简介
一、前言
提到CRISPR,大家耳熟能详的可能是麻省理工学院-哈佛大学Broad研究所、有着“CRISPR之父”之称的张锋,也可能是加州大学伯克利分校的女神科学家Jennifer Doudna。


- CRISPR全称是Clustered Regularly Interspaced Short Palindromic Repeats(成簇的规律间隔的短回文重复序列)。该序列于1987年被发现
- 而Cas的全称是CRISPR associated(CRISPR关联)
- 由于名字太长,后来大家都简称为CRISPR/Cas系统
CRISPR/Cas这项技术自从问世以来,已经吸引了无数欢呼和掌声,在短短两三年之内,它已经成为了生物科学领域最炙手可热的研究工具,但其实CRISPR/Cas系统早就存在自然界中了。CRISPR/Cas系统是一种原核生物的免疫防御系统,用来抵抗外来遗传物质的入侵,比如噬菌体病毒等。 同时,它为细菌提供了获得性免疫(类似于哺乳动物的二次免疫),当细菌遭受病毒入侵时,会产生相应的“记忆”。当病毒二次入侵时,CRISPR系统可以识别出外源DNA,并将它们切断,沉默外源基因的表达,抵抗病毒的干扰。是不是觉得和真核生物中RNA干扰(RNAi)的原理很相似?正是由于这种精确的靶向功能,CRISPR/Cas系统被开发成一种高效的基因编辑工具。在CRISPR/Cas系统中,CRISPR/Cas9系统是研究最深入,应用最成熟的一种类别。
CRISPR/Cas9是继锌指核酸内切酶(ZFN)”、“类转录激活因子效应物核酸酶(TALEN)”之后出现的第三代基因组定点编辑技术。
二、结构简介
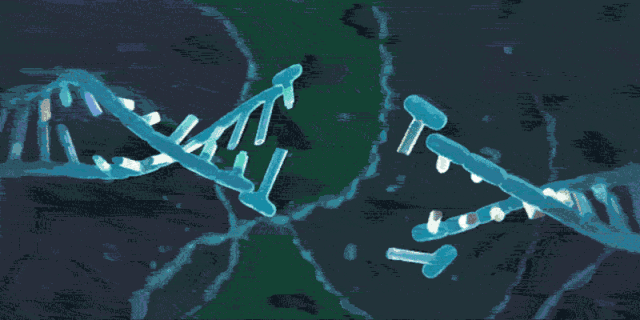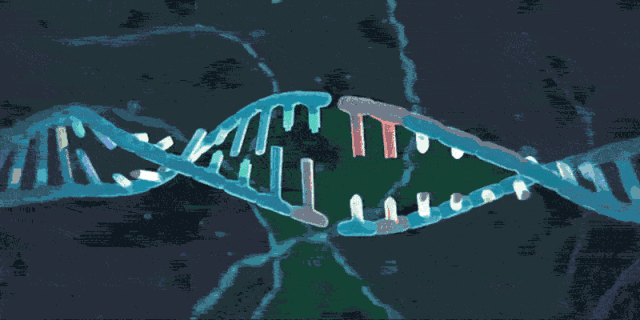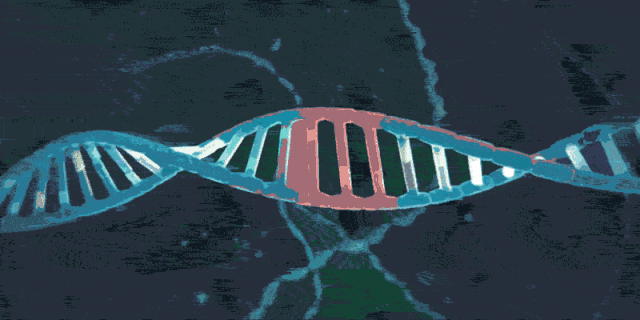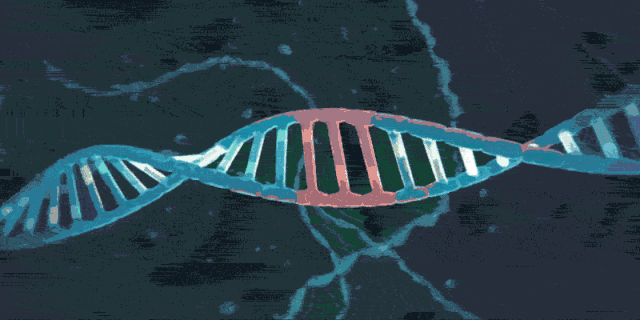
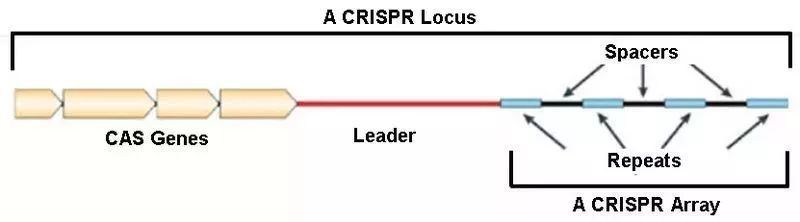
CRISPR簇是一个广泛存在于细菌和古生菌基因组中的特殊DNA重复序列家族,充当了防御外源遗传物质的“基因武器”,分布在40%的已测序细菌和90%的已测序古细菌当中。下图展示了完整的CRISPR位点(Locus)的结构。
其中,CRISPR序列由众多短而保守的重复序列区(Repeats)和间隔区(Spacers)组成。
- Repeats含有回文序列,可以形成发卡结构。
- 而Spacers比较特殊,它们是被细菌俘获的外源DNA序列。 这就相当于细菌免疫系统的“黑名单”,当这些外源遗传物质再次入侵时,CRISPR/Cas系统就会予以精确打击。
- 而在上游的前导区(Leader)被认为是CRISPR序列的启动子。
- 另外,在上游还有一个多态性的家族基因Cas,该基因编码的蛋白均可与CRISPR序列区域共同发生作用。Cas基因与CRISPR序列共同进化,形成了在细菌中高度保守的CRISPR/Cas系统。目前已经发现了Cas1-Cas10等多种类型的Cas基因。
cas酶简介
Makarova KS等人在2015年根据crRNA招募效应蛋白的组成将其分为两级,1级CRISPR系统利用多个Cas蛋白与crRNA形成效应复合物,2级CRISPR系统利用单个大Cas蛋白与crRNA形成效应复合物。自然组成越简单越可以获得更大的应用。那Cas9与Cpf1选择哪个更好呢?
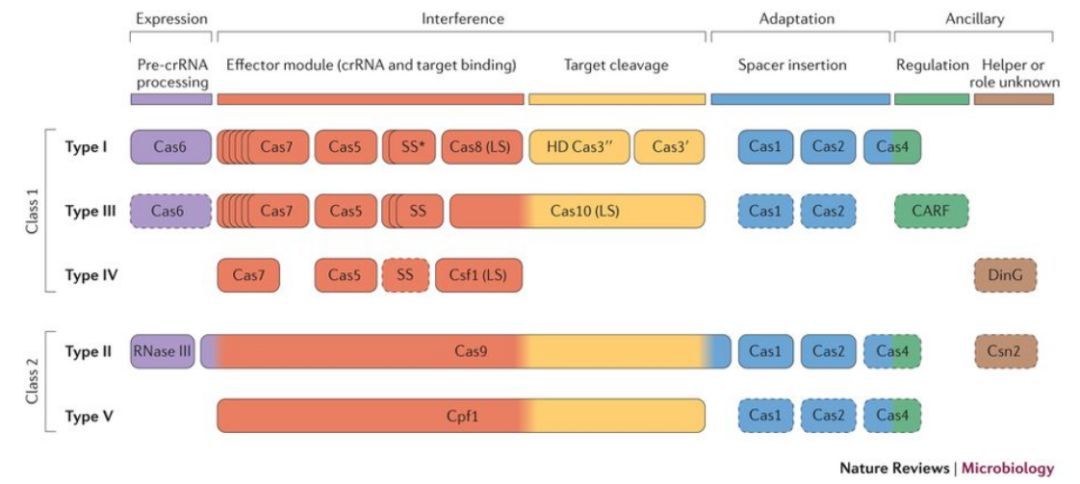
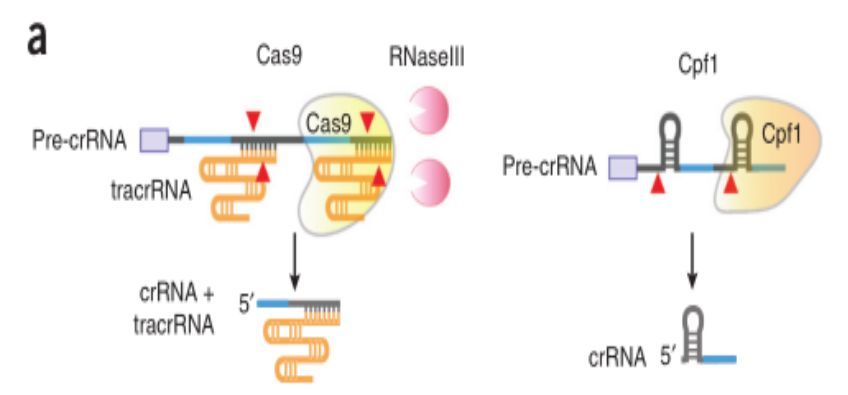
Cas(CRISPR-associated protein):CRISPR相关蛋白,是一种核酸内切酶。在细菌防卫的第三阶段,crRNA介导Cas核酶切割靶DNA,靶DNA必须包含一个物种特异性的PAM(protospacer adjacent motif )序列。不同的Cas蛋白识别不同的PAM序列,最常见的S. pyogenesCas9 (SpCas9) 识别的PAM序列是5’ NGG 3’,虽然NGG在人类基因组中平均每8个碱基出现一次,但还是有其限制性,由此衍生了很多SpCas9变种酶或者SpCa9直系同源酶如SaCas9。
Cpf1与Cas9除了识别的PAM位点不同外,其CRISPR系统也有几点不同。1) CRISPR-Cas9系统中成熟的crRNA会与tracrRNA部分碱基互补配对(crRNA和tracrRNA称为gRNA,有时会将其融合在一起称为sgRNA,single guide RNA)形成一个发夹结构招募Cas9蛋白,而CRISPR-Cpf1系统crRNA独自形成一个发夹结构招募Cpf1蛋白;2) Cas9-crRNA复合物切割靶DNA形成一个平末端的双链断裂缺口,而Cpf1-crRNA切割靶DNA形成一个黏性末端。这就提出问题,与Cas9系统相比,Cpf1系统不需要tracrRNA,而且黏性末端可能更有助于发生同源重组修复,是否Cpf1酶更好呢?
三、工作原理
在具体的工作过程中,CRISPR序列和Cas蛋白配合,大体上分3步来执行防御功能。

3.1 外源DNA俘获
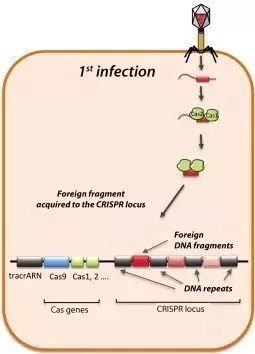
简单来说,CRISPR/Cas系统首先要取得一段外源DNA,实现“黑名单登记”。CRISPR/Cas系统将识别出入侵者(比如病毒)的“名字”(PAM)并找到它的“身份证”(原间隔序列),然后把入侵者身份信息作为“档案”(间隔序列)记录到“黑名单”(CRISPR序列)中。下图展示了第一阶段的工作原理。当噬菌体病毒首次入侵细菌,病毒的双链DNA被注入细胞内部。
- 直接重复序列(repeat)被易变的DNA序列间隔开组成CRISPR序列,此时的DNA序列称为space
- Cas蛋白切割外来DNA片段,外来DNA片段称为protospacer,然后将其整合到CRISPR序列上,这时外来DNA片段更名为spacer
- 原间隔序列向两端延伸的几个碱基都十分保守,被称为前间隔序列临近基序(protospacer adjacent motif,PAM)。PAM通常由NGG三个碱基构成(N为任意碱基)。
病毒入侵时,Cas1和Cas2编码的蛋白将扫描这段外源DNA,并识别出PAM区域,然后将临近PAM的DNA序列作为候选的前间隔序列。随后,Cas1/2蛋白复合物将前间隔序列从外源DNA中剪切下来,并在其他酶的协助下将原间隔序列插入临近CRISPR序列前导区的下游。然后,DNA会进行修复,将打开的双链缺口闭合。这样一来,一段新的间隔序列就被添加到了基因组的CRISPR序列之中。
3.2 crRNA合成
目前的研究表明,CRISPR/Cas系统共有三种方式(TypeⅠ、Ⅱ、Ⅲ)来合成crRNA,CRISPR/Cas9系统属于TypeⅡ(目前最成熟也是应用最广的类型)。当病毒入侵时,CRISPR序列会在前导区的调控下转录出pre-CRISPR-derived RNA (pre-crRNA)和trans-acting crRNA(tracrRNA)。其中,tracrRNA是由重复序列区转录而成的具有发卡结构的RNA,而pre-crRNA是由整个CRISPR序列转录而成的大型RNA分子。随后,pre-crRNA,tracrRNA以及Cas9编码的蛋白将会组装成一个复合体。它将根据入侵者的类型,选取对应的“身份证”(间隔序列RNA),并在核糖核酸酶Ⅲ(RNaseⅢ)的协助下对这段“身份证”进行剪切,最终形成一段短小的crRNA(包含单一种类的间隔序列RNA以及部分重复序列区)。crRNA,Cas9以及tracrRNA组成的最终的复合物,为下一步剪切做好准备。
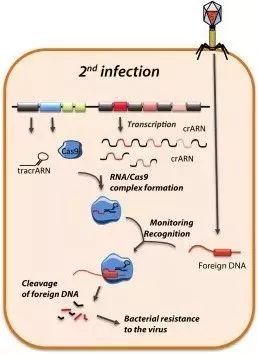
3.3 靶向干扰
如下图所示,在病毒的二次感染中,Cas9/tracrRNA/crRNA复合物可以对入侵者的DNA进行精确的打击。复合物会扫描整个外源DNA序列,并识别出与crRNA互补的前间隔序列。这时,复合物将定位到PAM/前间隔序列的区域,DNA双链将被解开。crRNA将与互补链杂交,而另一条链则保持游离状态。随后,Cas9蛋白发挥作用,剪切crRNA互补的DNA链和非互补的DNA链。最终,Cas9使双链断裂(DSB)形成,外源DNA的表达被沉默。
四、应用价值
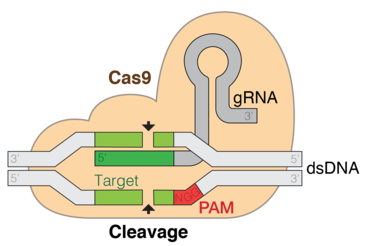
CRISPR/Cas的强大之处在于,其可以对基因进行定点的精确编辑。在向导RNA(guide RNA, gRNA)和Cas9蛋白的共同作用下,细胞基因组DNA(看成外源DNA)将被精确剪切。但是,被CRISPR/Cas9剪切需要满足几个条件:
- 待编辑的区域附近需要存在相对保守的PAM序列(NGG)。
- 向导RNA要与PAM上游的序列碱基互补配对。
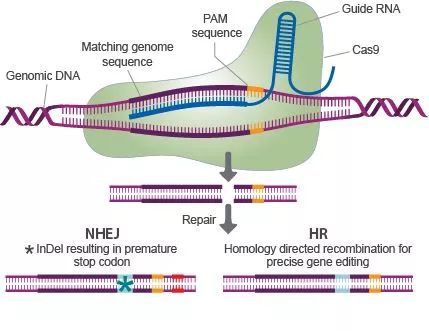
最基础的应用就是基因敲除。如果在基因的上下游各设计一条向导RNA(gRNA1,gRNA2),将其与含有Cas9蛋白编码基因的质粒一同转入细胞中,gRNA通过碱基互补配对可以靶向PAM附近的目标序列,Cas9蛋白会使该基因上下游的DNA双链断裂。随后生物体自身存在着DNA损伤修复的应答机制,会将断裂上下游两端的序列连接起来,从而实现了细胞中目标基因的敲除。如果在此基础上为细胞引入一个修复的模板质粒(供体DNA分子),这样细胞就会按照提供的模板在修复过程中引入片段插入(Knock-in)或定点突变(site-specific mutagenesis)。这样就可以实现基因的替换或者突变。随着研究的深入,CRISPR/Cas技术已经被广泛的应用,除了基因敲除,基因替换等基础编辑方式,它还可以被用于基因激活,疾病模型构建,甚至是基因治疗。
4.1 CRISPR:看魔剪如何征服DNA
首先,来看看曾经轰动一时的案例:中山大学研究人员利用CRISPR-Cas9校正人胚胎中的突变。中国科学家利用CRISPR/Cas9对人胚胎中会导致地中海贫血的β珠蛋白基因突变成功地进行修饰,目的是希望利用CRISPR对这种基因突变进行校正,以便实现利用基因疗法治疗地中海贫血。文章发表在《蛋白·细胞》(Protein & Cell)杂志上[1]。
β-地中海贫血是一种潜在地危及生命的血液疾病,它在全世界的患病率大约是十万分之一。研究人员利用来自一名β-地中海贫血患者的组织构建出克隆胚胎,随后对这些胚胎中的DNA进行了测序,发现一种单核苷酸错误,即应当为A的碱基被替换为了G。接下来他们研究利用CRISPR将这种碱基转换回去,即实现G到A。这也是首次证实利用CRISPR系统治愈人类胚胎中的遗传疾病是可行的。但这项研究造成巨大影响力的原因还有:它是在婴儿胚胎里面开展的,虽然医生丢弃的是不会成功孕育出婴儿的异常胚胎,但这点还是受到了部分学者的批评和攻击。
就在这不久之后,《新英格兰医学杂志》(NEJM)报道了巴塞尔大学研究人员利用CRISPR找到导致红细胞增多症的第一个遗传突变。[2]。
通过使用全基因组连锁分析和基因测序,研究人员发现所有受影响的家族成员的EPO基因中都缺少一个单碱基。而EPO的增加正是导致血红细胞过多产生的原因。但困惑的是,这个碱基的缺失导致了基因编码读码框架发生移动,最终导致EPO基因功能缺失而不是增强。但实际情况是,病人血液中EPO的含量是增加而不是降低。最终,还是CRISPR帮研究人员找到了答案。原来,EPO基因中还有一个隐藏的mRNA,正常情况下并不参与形成EPO。基因突变导致这个基因的读码框架发生移动,从而导致了这个基因产生了更多的EPO。
除了在血液方面的应用,CRISPR在发育方面也卓有建树。近期《自然》(Nature)就报道了英国和韩国的研究人员利用CRISPR/Cas9揭示OCT4基因在人胚胎早期发育中发挥着关键作用。在正常情形下,OCT4基因在人胚胎的最初几天发育中是有活性的,它驱动受精卵分裂,大约7天后形成一种有大约200个细胞组成的球体,即囊胚。在实验中,他们用CRISPR/Cas9阻断人胚胎OCT4的表达后,这些胚胎的发育都停止了[3]。
CRISPR另外一个价值就是能够用来追踪细胞,这个功能在癌症方面都有着重要的作用。斯坦福大学的研究人员就将CRISPR基因编辑技术同DNA条形码技术结合,有效追踪癌症进展。
人类的癌症并不仅仅只有一种肿瘤抑制突变,其存在多种突变组合。为了了解不同的突变基因是如何相互作用的,研究人员们耗费了数年的努力来绘制图谱,包括构建多种不同谱系的遗传修饰化小鼠,其中每一种都携带不同的失活肿瘤抑制基因。而如果想探索所有的可能性组合,研究人员就需要上千只小鼠。
CRISPR-Cas9的强大之处在于,可以轻易地替换、修改或删除生物体内的基因序列,从而在单个小鼠的肺中创造出多种基因不同的肿瘤。问题在于,为了得出关于不同基因突变组合效应的有用结论,科学家需要一种精确的方法来标记追踪不同肿瘤的生长。但如果将CRISPR-Cas9和DNA条形码技术(利用生物体DNA中一段保守片段对物种进行快速准确鉴定的技术)相结合以此来追踪癌症的生长发展情况,就能帮助科学家们在实验室中复制出癌症患者机体中所观察到的遗传多样性。就比如将短的、独特的DNA序列(DNA条形码)黏附于小鼠肺内的单个肿瘤细胞中,这样每一个序列的功能就像是遗传条码,当每个癌细胞扩增时,条码的数量会随之增加。最后只需要将整个癌肺取出,然后用高通量的DNA测序和计算来分析条形码出现的频率,从而精确的确定肿瘤的大小。
也就是说,研究人员可以在同一个小鼠身上产生大量具有特定遗传特征的肿瘤,并在规模和精度上分别跟踪它们的生长。研究人员最终只在几个月之内就完成了相关实验,只用了不到24只小鼠。结果发表在《自然·遗传学》(Nature Genetics)上[4]。
无独有偶,Nature重量级子刊《自然·生物科技》(Nature Biotechnology)也报道了德国研究人员利用CRISPR/Cas9诱导的DNA瘢痕序列追踪细胞谱系[5]。
在生物界一直有一个问题困着大家,功能不同的细胞系到底来自于何处?而CRISPR有一个特点是总在确切的位点上进行切割,受此启发,德国科学家团队开发出一种被称作LINNAEUS(lineage tracing by nuclease-activated editing of ubiquitoussequence, 通过对普遍存在的序列进行核酸酶激活的编辑来开展谱系追踪)的技术,它能够让人们确定细胞类型和每个细胞的谱系。
在斑马鱼胚胎细胞中如何切割DNA,在下一次细胞分裂发生之前,给DNA修复时间不超过15分钟。修复工作必须快速完成,这也是错误累积的地方,这种错误序列我们就称之为DNA瘢痕序列。DNA中的瘢痕序列具有随机的长度,而且它们的确切位置也会发生变化。子细胞在细胞分裂过程中遗传这些瘢痕序列。当这些瘢痕序列汇总在一起时就像条形码一样发挥作用,能够确定每个细胞的谱系。
因此研究人员设计了一个实验,在斑马鱼的胚胎内注入CRISPR-Cas9系统,这些斑马鱼事先被转入了红色荧光蛋白(RFP)。在接下来的时间内,Cas9重复性地切割斑马鱼体内的RFP。这些斑马鱼胚胎中的红色荧光逐渐消失的同时,数千个瘢痕序列在细胞中DNA损伤区域中形成。最终通过分析瘢痕序列,就能确定细胞是来自于哪个祖细胞。
综上所述,CRISPR在DNA 编辑方面起着重要作用,也衍生出众多的新技术。
4.2 当CRISPR遇上CAR-T:看强强如何联手攻克癌症
通用型CAR-T
首先让我们来聊聊通用型CAR-T。目前,市场上大部分的CAR-T细胞是利用患者自身的T细胞来生产的。这个过程不仅耗时耗力,价格昂贵,而且还受限于当前的生产制造能力。如果科学家们能够想办法生成通用型的CAR-T细胞,那这类疗法将有望变得更便捷、更便宜。因为,这些现成的(off-the-shelf)细胞将增加能够接受单一CAR-T细胞产品治疗的患者数量。
https://www.nature.com/articles/nrd.2016.238
但有两个问题成为通用型CAR-T的主要障碍,移植物抗宿主病(graft-versus-host disease,GVHD)和宿主排斥。不过幸运的是,美国纪念斯隆凯特林癌症中心科学家领导的研究小组发现,同种异体供体CD19特异性的CAR-T细胞在杀伤肿瘤细胞的同时,所导致的GVHD发生的风险是最少的。文章发表在《Nature Medicine》上。
https://www.nature.com/articles/nm.4258
基于这样一个天然的优势,研究人员就想着利用CRISPR对CAR-T进行基因编辑,降低CAR-T的抗原性。CAR-T领域的先驱者Carl June教授就在《Clinical Cancer Research》杂志上发表文章证实,体外和动物模型研究中,用CIRSPR敲除TCR(T cell receptor)和B2M(β-2 microglobulin)这两个和免疫排除有关的基因后,T细胞同种异体反应性(alloreactivity)降低,且没有导致GVHD。
http://clincancerres.aacrjournals.org/content/23/9/2255.long。
无独有偶,中国和英国科学家也发现,用CRISPR敲除TRAC (T cell receptor alpha constant chain)后可以构建的通用型CAR-T。
https://www.ncbi.nlm.nih.gov/pubmed/29605708
https://www.nature.com/articles/cr2016142
笔者在此还想补充的一点是,通用型CAR-T还有另外一种解读,就是同一种T细胞可以加载不同的抗原。这个概念是由MIT和波士顿大学的科学家提出来的。如下图所示,在T细胞上表达通用型的受体,然后这个通用型的受体可以连接不同的靶点抗原,开发出一种分离的、通用、可编程式CAR 系统用于T细胞治疗,他们起名为SUPER CAR-T。也就是说,患有不同癌症的患者只需要输入同一种SUPER CAR-T,然后再输入相应的抗原(比如肺癌特异性抗原或者淋巴瘤特异性抗原等),这大大加快了治疗进程。
https://www.cell.com/cell/fulltext/S0092-8674(18)30362-3
增强型CAR-T
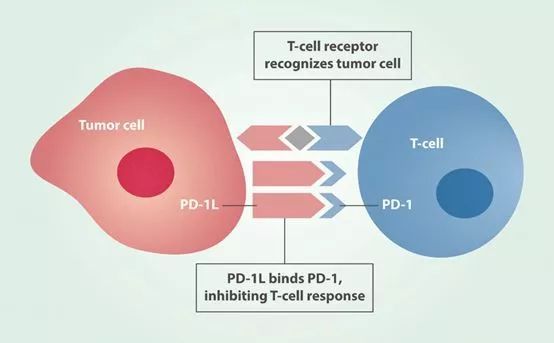
CRISPR除了用于产生通用型CAR-T,还能通过敲除免疫共抑制通路或信号分子的基因(如CTLA4、PD1)提高CAR-T细胞的功效。比较有代表性的就是NIH(美国国立卫生研究院)下属Recombinant DNA Advisory Committee批准的由Carl June教授领导的一项CRISPR临床试验。在该试验中,研究人员将利用CRISPR/Cas9在靶向黑色素瘤的CAR-T中敲除编码PD-1的基因以及内源性T细胞受体的基因。此项研究由宾夕法尼亚大学和它的合作伙伴—加州大学旧金山分校以及MD安德森癌症中心共同负责。《Nature Biotechnology》杂志还特意报道了该研究团队计划在I期临床试验结果。敲除PD-1之后,结果表明CAR-T的杀伤性更强。
https://www.nature.com/articles/nbt0117-3
但抑制PD-1通路也不是万能的,《Nature》杂志报道,如果在T细胞非霍奇金淋巴瘤(T-NHL)的癌症小鼠模型中使用PD-1抑制剂免疫疗法,非但没发挥抗癌作用,而且还促进了肿瘤的进展。这个结果其实是在意料之中,因为在T-NHL中,T细胞本就是肿瘤细胞,抑制了PD-1通路只会加速肿瘤生长。既然PD-1/PD-L1不行,那CAR-T能行吗?这时又出现另外一个问题:CAR-T细胞和癌性T细胞之间目标抗原的共同表达会导致CAR-T细胞的自相毁灭。这些问题都限制了CAR-T细胞疗法的发展。
不过不用担心,来自圣路易斯华盛顿大学医学院的科学家们利用基因编辑技术CRISPR对人类T细胞进行了改造,去除了CD7 和T细胞受体α链(TRAC)表达,这是一种针对CD7+ T细胞恶性肿瘤,避免了“自相残杀”的CAR-T细胞疗法(UCART7)。其能够在保护正常T细胞的情况下,对癌性T细胞发起攻击。研究结果于近日发表在权威学术期刊《Nature》子刊《Leukemia》上,结果显示:接受基因编辑改造的以CD7为靶点的T细胞治疗组的小鼠,中位生存期为65天,而接受对照组小鼠的中位生存期仅有31天。
https://www.nature.com/articles/s41375-018-0065-5
总结一下,CRISPR联合CAR-T确实能到起到1+1大于2的效果,一起强强联手消灭癌症。希望未来能有更多的突破出现。
深度剖析CRISPR的临床应用和市场前景
4.3 临床应用
说到临床应用,中国科学家在这方面拔得头筹。虽然张锋、Jennifer Doudna和Carl June等大牛一直在争先恐后的申请临床试验,但这一领域的格局或许要被一批“默默无闻”的中国科学家改写了!就在2016年7月,《Nature》宣布,在获得了医院审查委员会的伦理批准之后,四川大学华西医院肿瘤学家卢铀教授将开展全球首个CRISPR人体试验,利用CRISPR技术编辑的T细胞,来治疗化疗、放疗以及其它疗法治疗无效的转移性非小细胞肺癌患者这一临床试验。事实上,中国在CRISPR领域一直行动很快,已经创造了多个第一,如第一个CRISPR编辑人类胚胎,第一个CRISPR编辑猴子。
试验具体操作过程就是,首先从患者的血液中提取出T细胞,然后用CRISPR/Cas9技术敲除PD-1基因。该基因是人体免疫反应的关键开关,删除它能够恢复T细胞的抗肿瘤能力。经CRISPR编辑后的T细胞在实验室中扩增后,再回输到患者血液中。当经过改造的T细胞循环到肺癌组织中时,就能不受肿瘤细胞抑制并将其消灭。这个方案还有一个值得改进的地方,就是研究小组可以选择从肿瘤部分获取T细胞,因为这些T细胞已准备好特异性攻击癌症。
当然除了中国,其他国家也在这个领域投入了大量精力。除了在前期文章“当CRISPR遇上CAR-T:看强强如何联手攻克癌症”中提及的Carl June领导的利用CRISPR/Cas9在CAR-T中敲除编码PD-1的基因以及内源性T细胞受体的基因的研究以外,美国的CRISPR Therapeutics公司(第一家获欧洲监管机构许可开展临床试验的公司)就在积极开展利用CRISPR修复遗传性血液病β-地中海贫血患者的基因缺陷。
该公司今年4月份已经向FDA提交了镰状细胞贫血病治疗药物 CTX001的临床试验申请。CTX001是 CRISPR Therapeutics 和 Vertex 制药公司共同开发、共同商业化的一种研究性体外基因编辑疗法。其通过编辑患者的造血干细胞基因,来提高红细胞中的胎儿血红蛋白(HbF)水平,从而实现β-地中海贫血和镰状细胞病的治疗。HbF是婴孩出生时天然存在的携氧血红蛋白的一种形式,后来逐渐被成人形式的血红蛋白代替。通过升高HbF,CTX001 可能会缓解β-地中海贫血患者的输血需求以及镰状细胞患者的疼痛和虚弱症状。虽然因为审查原因FDA暂停了关于CTX001的研究性新药申请(IND),但在欧洲的临床 I 期和Ⅱ期试验计划仍然不变,预计将在 2018 年下半年开始试验。
此外,Editas公司也将开展针对遗传性视网膜疾病:10型先天性利伯氏黑蒙(Lebercongenital amaurosis type 10,生物通译)的I期试验,这一研究将使用gRNA-Cas9核酸酶,纠正编码中心体蛋白290(CEP290)的基因突变。
4.4 市场分析
谈及CRISPR的市场,可以用炙手可热这个词来形容。前不久,张锋和David Liu共同建立的Beam The rapeutics成功刷屏,可以反应出市场的火热度。笔者在此不想赘述CRISPR的市场价值,而是想客观分析一下CRISPR的副作用。
首先吸引大家眼球的肯定是CRISPR的脱靶效应,就是本来想编辑目标基因,结果“子弹”跑偏了,编辑了一个无关基因。最具代表性的就是《Nature Methods》的一篇文章[1]声称来自哥伦比亚大学等机构的研究人员发现,在治愈小鼠相关遗传性疾病的同时,基因编辑常用的工具 CRISPR/Cas9 可能会造成很多意料之外的基因突变。该研究的发表一度造成了一大波学界关于CRISPR技术的批评和指责,并影响了数个正在开展的基于CRISPR 基因编辑技术的临床试验。
不过很快就被其他科学家反驳,他们表示该研究在方法和逻辑上存在缺陷,其中包括样本数量太少,结论与数据不符合等。作为 CRISPR 领域的领军人物,Editas Medicine 的张锋团队和Intellia Therapeutics 的 Jennifer Doudna团队的科学家们也在当时分别写信给《Nature Methods》编辑部,认为该论文的结论存在偏颇,并要求将该论文撤稿。
面对质疑,《Nature Methods》展开了调查,并最终作出撤稿的决定。其给出撤稿的理由是,文章中所采用的小鼠虽然来源于同一“祖先”,然而,基因编辑的小鼠和对照小鼠也并非属于亲兄弟姐妹的关系,即便是亲兄弟姐妹,其基因组本身存在的突变也可能有很多不同。遗传背景来源不清楚,这就造成一个巨大问题:这些小鼠在采用CRISPR基因编辑技术之前,其基因组本来就已经存在许多突变了,而非基因编辑之后造成的。其次,其他研究团队经过分析之后,也没有证据显示CRISPR基因编辑之后会造成突变。为了给这场争辩画上一个完美的句号,《Nature Methods》在3月30日发表来自基因编辑领域数十位顶尖学者和机构的多篇通讯文章,来回应这篇文章。
一波未平,后浪再起。就在前不久,两篇发表在《Nature Medicine》上的文章[2,3]再次把矛头指向CRISPR,有媒体报道称“使用CRISPR-Cas9成功编辑的多能干细胞很可能有p53 缺陷,这将增加肿瘤风险”,“这些基因编辑细胞就仿佛是定时炸弹一样”。不过这次,笔者得强调一下,我们得对这两篇文章进行正确的解读。
在此,笔者想引述一下知识分子对此事件回复的专题文章[4]。在第一篇文章[2]中,作者得出的结论是CRISPR-Cas9会触发p53基因介导的DNA损伤修复机制,这会使基因编辑变得更加困难,而如果人为去除p53基因,解除这种修复机制,会让细胞变得更容易被编辑。而在第二篇文章[3]中,作者觉得,如果p53功能健全,多数细胞会编辑失败。据他们估计,在多能干细胞中,p53使CRISPR的效率降低了17倍之多。所以实际上,这两篇文章的结论是p53缺陷的细胞更容易被编辑,而并不是细胞被编辑后更容易造成p53的缺陷。目前还无法证实CRISPR有致癌风险。
当然,两篇文章也引出了一个令人担忧的假想:CRISPR会不会倾向于编辑含有p53缺陷的细胞,而丧失了这个“基因守护者”的细胞会有很高的致癌风险。但多位专家认为,p53缺陷的细胞更容易被编辑并不一定意味着编辑后细胞更容易出现p53的缺陷。CRISPR只是导致DNA双链断裂的其中一种方式,用CRISPR来产生DNA断裂,并不比其他方式产生的断裂特殊,更不会特别去影响p53。
由此可见,真金不怕火来炼呀。任何技术都不是完美的,CRISPR也不例外。但CRISPR货真价实的基因编辑效率,让人不得不遐想其无限的未来。
行文至此,关于CRISPR的系列报道就告一段落,希望大家都能有所收获。
参考资料
