【9.1】mRNA cap
一种快速且经济高效地大规模生产 5' 加帽 RNA 的通用方法
真核 mRNA 5' 帽结构对于前体 mRNA 加工、mRNA 输出、翻译起始和 mRNA 稳定性是必不可少的
5’鸟嘌呤-N 7-甲基帽结构是真核mRNA的中心结构特征(Reddy et al. 1974 ; Furuichi and Miura 1975 ; Shatkin 1976)。帽结构的存在对于 mRNA 生命周期中的几乎所有后续步骤都是必不可少的,包括:
- 前 mRNA 剪接(Izaurralde 等人,1994 年;Fresco 和 Buratowski 1996 年;Schwer 和 Shuman 1996 年)、mRNA 输出(Hamm 和 Mattaj) 1990 年),以及翻译的开始(Both 等人 1975 年;Filipowicz 1978 年)。
- 此外,mRNA 帽结构通过保护 mRNA 免受 5'-3' 核酸外切降解(Furuichi et al. 1977)来稳定 mRNA ,并且帽结构的去除不可逆地针对转录物进行衰减(Parker and Song 2004)。
- 此外,获得帽结构是病毒确保其 RNA 有效翻译和逃避宿主免疫系统的常见机制(Daffis 等人,2010 年;Decroly 等人,2012 年)。
在基本的真核帽结构(m 7 G 或 cap-0)中,一个 5' N 7-甲基鸟苷通过一个不寻常的 5'-5' 三磷酸键连接到 mRNA 体上(图1A)。加帽酶通过羧基末端结构域 (CTD) 被募集到 RNA 聚合酶 II ( Cho et al. 1997 ) 并在转录物达到约 20 到 30 个核苷酸 (nt) 的长度时进行共转录(Salditt-Georgieff 等1980 年)。

只罗列部分实验结果和观点
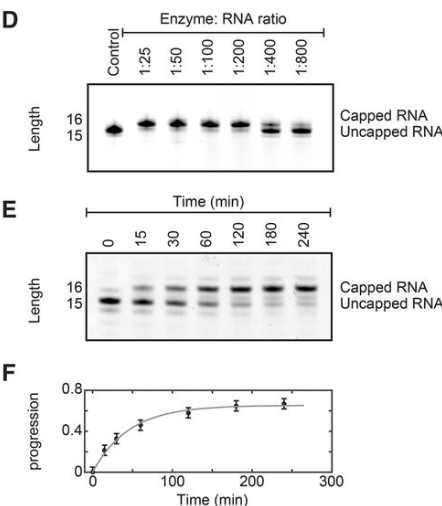
一、加冒率反应条件
加冒率取决于
- 酶:RNA 的比例,在所有后续实验中,我们在进行大规模加帽反应之前使用小规模测试来优化该比例。我们发现最佳酶:底物比例在 1:10 和 1:500 之间变化,具体取决于 RNA 体。
- 更长的孵育时间可以增加加帽反应的产率(图 1 E);然而,酶会在数小时内失去活性(图 1F)。因此,在不超过 1-2 小时的反应时间内最可靠地确保完全封盖。
二、序列对加帽的影响
为了测试牛痘加帽酶的效率对 RNA 体的长度、序列和二级结构的依赖性,我们进行了大量加帽反应,其中我们改变了单个参数。为了测试底物长度如何调节加帽效率,我们使用 RNA 体在 2 到 100 nt 之间的底物进行了加帽反应。如果 RNA 体超过 40 nt,则无法使用标准的尿素-PAGE 分析直接跟踪加帽反应,因为一个碱基的差异无法再清楚地解决。因此,为了跟踪加帽反应,我们设计了一种底物,该底物包含一个独特的 RNase A 切割位点,该位点位于 RNA 主体 5' 末端下游 15 个碱基处(图 2 A)。通过这种方式,我们可以对全长完整 RNA 进行加帽反应,并观察用 RNase A 切割后加帽反应的进程。重要的是,从这组实验中我们可以得出结论,40 到 100 nt 之间的 RNA 是完全并有效地封顶。为了测试非常短的 RNA 是否也是痘苗病毒加帽酶的底物,我们使用了 2 到 5 nt 之间的 RNA。有趣的是,我们观察到加帽酶甚至可以有效地完全加帽二核苷酸序列(图2B )。总之,牛痘加帽酶能够有效地加帽 RNA,完全独立于底物长度。
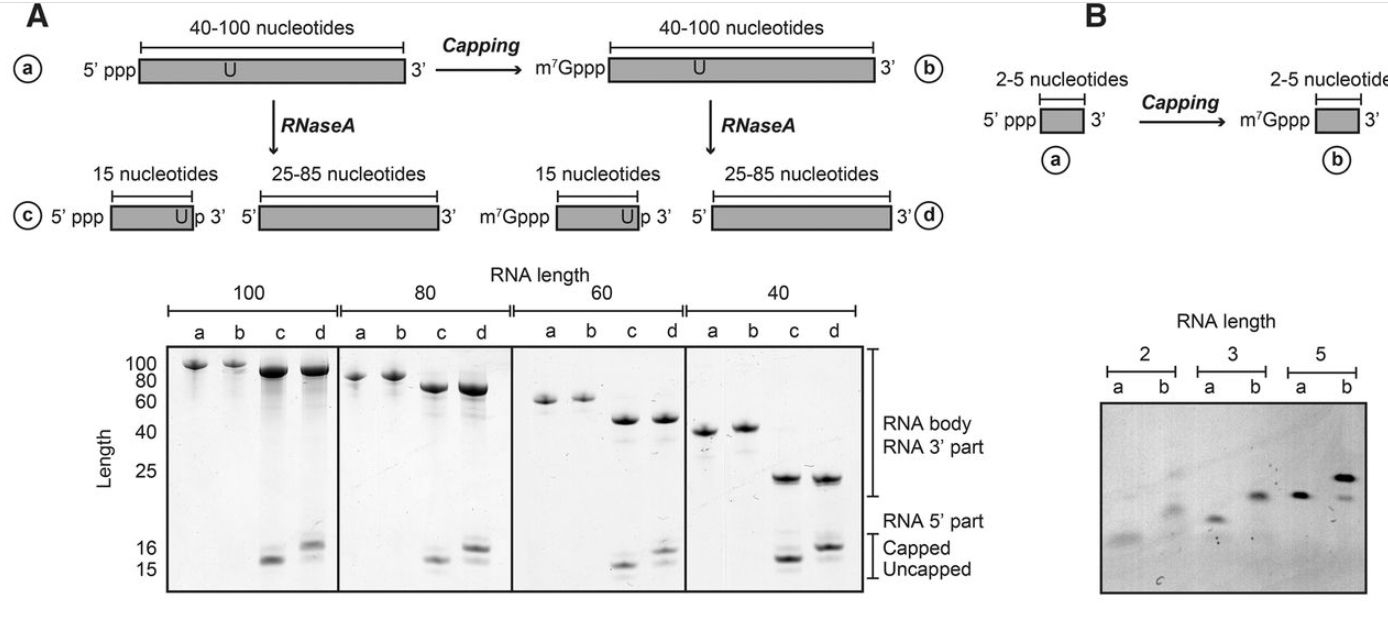
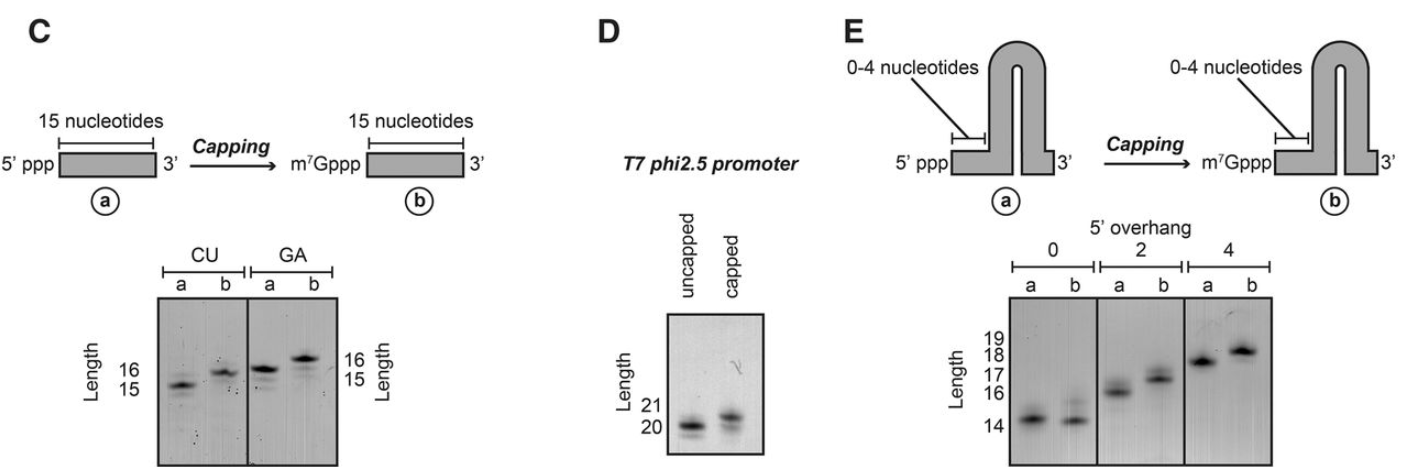
RNA 加帽反应稳健且独立于底物 RNA 长度、序列或二级结构。( A ) 使用不同长度(100 到 40 nt)的 RNA 底物进行加帽反应。为了可视化加帽效率,在位置 15 处引入了单个 U。RNA 酶 A 消化 RNA(加帽前后)释放仅包含 15 nt 的底物的 5' 端。使用尿素-PAGE 分析可以很容易地观察到由于封端反应引起的质量转移。所有 RNA 的加帽效率接近 100%。( B ) 较短的 RNA 序列(2-5 个碱基)也可以使用纯化的加帽复合物有效加帽。( C) 加帽反应与序列无关,因为富含 GA 和 CU 的序列都可以以接近 100% 的效率加帽。请注意,由于碱基组成,GA 和 CU RNA 的迁移略有不同。( D ) 当 RNA 体的第一个碱基是腺嘌呤时,加帽反应是有效的。( E ) RNA 二级结构仅在没有 5' 突出端时干扰加帽反应。不含 5' 突出端的稳定 RNA 发夹不被加帽,而具有 2 或 4 个碱基的 5' 突出端的稳定发夹被有效加帽。
为了评估 RNA 的序列是否在加帽过程中起作用,我们使用了富含 GA 或富含 CU 的 RNA 体。两种底物都被完全有效地加帽,表明 RNA 主体的一级序列不影响加帽效率(图2C )。此外,我们确定当转录物的第一个碱基是腺嘌呤时,RNA 加帽反应也起作用,因为用 T7 φ2.5 启动子产生的转录物也被完全加帽(图2D)。
最后,为了测试 RNA 二级结构是否对加帽效率有影响,我们使用了一个非常稳定的 GC 发夹结构,在 5' 末端没有、两个或四个碱基突出。在这些 RNA 体中,只有包含完全碱基配对的 5' 末端的 RNA 发夹不是加帽酶的底物(图2E)。其他 RNA 体被有效且完全加帽,表明稳定的二级结构元件不会干扰加帽反应,除非 5' 端是稳定双链的一部分。
总之,我们在这里表明牛痘加帽酶能够以有效的方式加帽 RNA,这实际上独立于 RNA 体的长度、序列和结构。
加帽 RNA 的翻译和去帽分析
为了证明加帽 RNA 的功能,我们制备了一个编码 GFP 蛋白的加帽转录本,并将其用于体外翻译试验(补充图 S4)。正如预期的那样,加帽反应后翻译效率显着提高,证明修改正确发生。
图S4
参考资料
