【9.3.5】聚腺苷酸化信号(PAS,polyA信号,SV40)
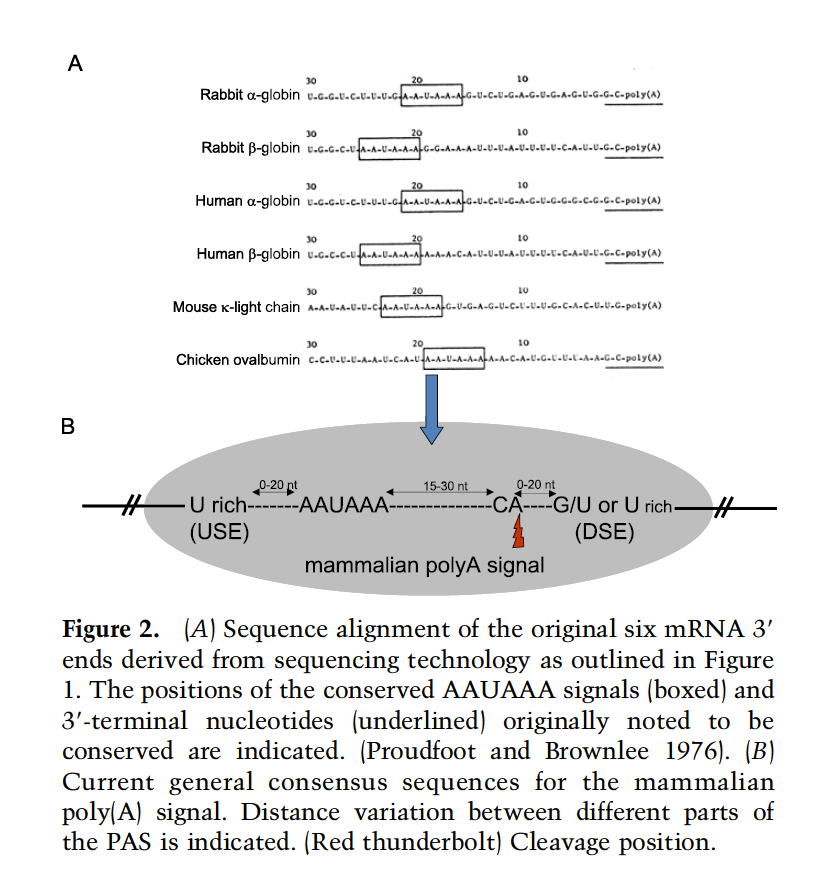
聚腺苷酸化信号 (PAS,Polyadenylation [poly(A)] signals) 是真核蛋白质编码基因的定义特征。核心序列基序 AAUAAA 在 1970 年代中期被鉴定,随后证明需要侧翼辅助元件来进行 3' 端切割和前信使 RNA(pre-mRNA)的多聚腺苷酸化以及促进下游转录终止。最近的基因组分析已经确定了 PAS 对真核 mRNA 的普遍性
在这两种情况下,突变 PAS 的亚克隆表明它失去了 poly(A) 形成功能。
尽管 AAUAAA 序列被证明是 mRNA 3'-末端多聚腺苷酸化所必需的,但其他序列元素被证明是完全重建功能性 PAS 所必需的。
- 特别是,在紧邻基因 3' 侧翼区域中的 mRNA 3' 末端之后存在的富含 GU 的序列(或下游序列元件,DS,downstream sequence element)被证明可以增强 3' 末端的形成(Gil and Proudfoot 1984 , 1987 ; McLauchlan等人,1985 年)。
- 类似地,紧邻 AAUAAA 上游的序列(上游序列元件,USE,upstream sequence element)在某些情况下可以作为 3' 端处理效率的增强元素。
- 最后,3’端切割位点的实际核苷酸也会影响这个过程的效率(Chen et al. 1995)。这被证明与人类凝血酶原基因具有医学相关性,该基因通常在切割位置的 5' 端携带一个 CG 二核苷酸序列。在大约 1% 的高加索人(包括我自己)中,该序列突变为 CA,导致轻度血栓形成倾向。这是由于获得更有效的 PAS 后凝血酶原基因表达增加了两倍(Gehring 等人,2001 年;Danckwardt 等人,2008 年)。
然而,其他真核模型系统揭示了 USE-AAUAAA-DSE 的哺乳动物模式(图 2B) 在真核生物中通常是保守的,即使在出芽酵母中,这些顺式元件的更大变化似乎是可以容忍的(Graber 等人,1999 年)。PAS 的一个显着特征是,根据 AAUAAA 和 DSE 信号的准确定位和定义方式,poly(A) 添加的实际位点可能会因数个到数十个核苷酸而异(Zhang 等人,1986 年;Sheets 等人,2006 年)。 1990 年;田等人,2005 年)。这在酵母 PAS 中尤为明显,这可能是由于序列保守性较低,不能准确定义多聚腺苷酸化的位置(Zhao et al. 1999)。这与前体 mRNA 剪接形成鲜明对比,前体 mRNA 剪接必须始终发生在精确的核苷酸位置,以保持 mRNA 的正确阅读框进行翻译。
常见的PAS
- tk polyA, SV40 late and early polyA or BGH polyA
- mammalian terminators (SV40, hGH, BGH, and rbGlob)
SV40 PolyA(猴空泡病毒PolyA, 简称PolyA)序列是有转录终止作用和使转录的mRNA 添加PolyA 尾的 DNA 序列(240 bp), 含有AATAAA 六核苷酸多腺苷化信号(Polyadenylation signal)。
AATAAA六核苷酸是最早被认识到的mRNA多腺苷化信号[1], 90%的真核mRNA 的AATAAA 序列都在距剪切位点的30 个核苷酸的范围内[2], 它指导核酸内切酶在此序列下游15~30 碱基处的特定位点上裂解前体mRNA, 然后通过多聚腺苷酸聚合酶的催化作用, 在3′-OH 上逐一引入100~300 个腺嘌呤核苷酸。AATAAA 六核苷酸已经显示出其分子和生理的关联性:例如, 一大类导致地中海贫血的血红蛋白病都归因于这个序列的点突变[3,4]。人基因中多腺苷酸化信号不是严格保守的, 单碱基变异发生率很高, 通过EST 定位研究推定mRNA 3′端仅73%具有明显六核苷酸多腺苷酸化信号[5]。

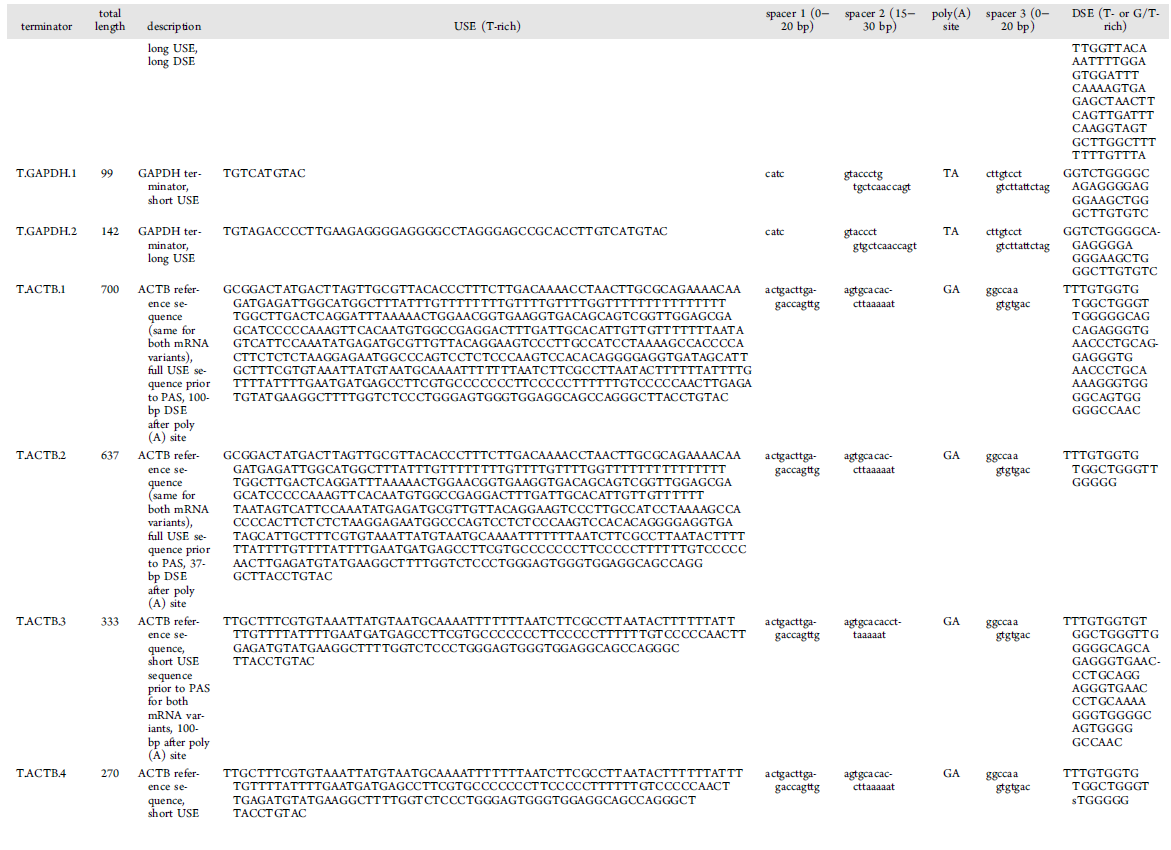
SV40 poly(A) signal:
TAAGATACATTGATGAGTTTGGACAAACCACAACTAGAATGCAGTGAAAAAAATGCTTTATTTGTGAAATTTGTGATGCTATTGCTTTATTTGTAACCATTATAAGCTGCAATAAACAAGTT
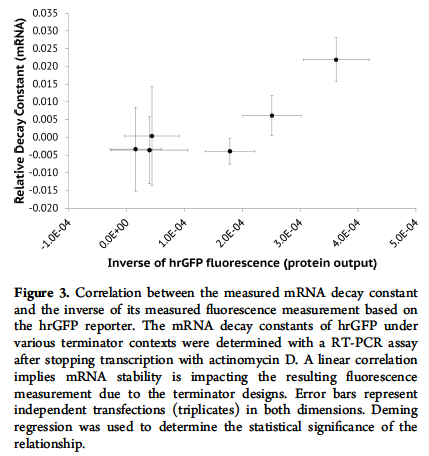
终止子通过增加 mRNA 稳定性发挥作用
参考资料
- Design and Evaluation of Synthetic Terminators for Regulating Mammalian Cell Transgene Expression 。 https://pubs.acs.org/doi/10.1021/acssynbio.8b00285#
- 2012,SV40 PolyA 序列及其AATAAA 信号对上游GFP 基因表达及转录终止的影响
- 2019, Design and Evaluation of Synthetic Terminators for Regulating Mammalian Cell Transgene Expression 。 https://pubs.acs.org/doi/10.1021/acssynbio.8b00285#
